Пути образования и использования ацетил-КоА. Биосинтез и значение кетоновых тел. Пределы изменений концентрации кетоновых тел в крови в норме, при голодании и сахарном диабете.
АцетилКоА- это центральный метаболит липидного обмена.
Источники: 1)Глюкоза 2)глицерин 3)АК> (при кратковременной напряженной мышечной работе) 4)Жирные к-ты (в-окисление при длительной мышечной работе, голодании, на холоде, при беременности и сахарном диабете). Пути использования образовавшегося ацетилКоА зависят от функционального состояния клетки (энергетический заряд) и ее специфики. Если в кл достаточно АТФ, то он используется на синтез ж к-т,т.к. АТФ активирует ключевой фермент ситеза ж к-т, а их накопление стимулирует синтез жира. Распад жира тормозится и в-окисление при этом тоже тормозится. Напряженная мышечная работа, стресс, увел-ие секреции катехоламинов активирует липолиз, в-окисление ж к-т; в этом случае актив-ся синтез кетоновых тел и ЦТК.
Пути использования: 1. окисляется в цикле Кребса(90%); 2. используется в синтезе ЖК (9%) 3. Образование В-гидрокси-в-метилглутарилКоА (а из него либо холестерин, либо кетоновые тела -1%)
При состояниях, сопровождающихся снижением глюкозы крови, клетки органов и тканей испытывают энергетический голод. Так как окисление жирных кислот процесс «трудоемкий», а нервная ткань вообще неспособна окислять жирные кислоты, то печень облегчает использование этих кислот тканями, заранее окисляя их до уксусной кислоты и переводя последнюю в транспортную форму кетоновые тела. К кетоновым телам относят три соединения близкой структуры ацетоацетат, 3-гидроксибутират и ацетон. Стимулом для образования кетоновых тел служит поступление большого количества жирных кислот в печень. Как уже указывалось, при состояниях, активирующих липолиз в жировой ткани, не менее 30% образованных жирных кислот задерживаются печенью. К таким состояниям относится голодание, сахарный диабет I типа, длительные физические нагрузки. Так как синтез ТАГ в этих условиях невозможен, то жирные кислоты из цитозоля попадают в митохондрии и окисляются с образованием кетонов. Кроме отмеченных ситуаций, количество кетоновых тел в крови возрастает при алкогольном отравлении и потреблении жирной пищи. При богатой жирами диете, особенно у детей, жирные кислоты не успевают включиться в состав ТАГ и ЛПОНП и частично переходят в митохондрии, что увеличивает синтез кетоновых тел. При алкогольном отравлении субстратом для синтеза кетонов является ацетил-SКоА, синтезируемый при обезвреживании этанола. В обычных условиях синтез кетоновых тел также идет, хотя в гораздо меньшем количестве. Для этого используются как жирные кислоты, так и безазотистые остатки кетогенных и смешанных аминокислот. Синтез ацетоацетата происходит только в митохондриях печени, далее он либо восстанавливается до 3-гидроксибутирата, либо спонтанно декарбоксилируется до ацетона. Далее все три соединения поступают в кровь и разносятся по тканям. Ацетон, как летучее вещество, легко удаляется с выдыхаемым воздухом и потом. Все кетоновые тела могут выделяться с мочой. Используются кетоновые тела клетками всех тканей, кроме печени и эритроцитов. Особенно активно, даже в норме, они потребляются миокардом и корковым слоем надпочечников. Реакции утилизации кетоновых тел примерно совпадают с обратным направлением реакций синтеза. В цитозоле 3-гидроксибутират окисляется, образующийся ацетоацетат проникает в митохондрии, активируется за счет сукцинил-SКоА и превращается в ацетил-SКоА, который сгорает в ЦТК.
Регуляция синтеза кетоновых тел. Регуляторный фермент синтеза кетоновых тел — ГМГ-КоА синтаза.
*ГМГ-КоА-синтаза — индуцируемый фермент; его синтез увеличивается при повышении концентрации жирных кислот в крови. Концентрация жирных кислот в крови увеличивается при мобилизации жиров из жировой ткани под действием глюкагона, адреналина, т.е. при голодании или физической работе.
*ГМГ-КоА-синтаза ингибируется высокими концентрациями свободного кофермента А.
*Когда поступление жирных кислот в клетки печени увеличивается, КоА связывается с ними, концентрация свободного КоА снижается, и фермент становится активным.
*Если поступление жирных кислот в клетки печени уменьшается, то, соответственно, увеличивается концентрация свободного КоА, ингибирующего фермент. Следовательно, скорость синтеза кетоновых тел в печени зависит от поступления жирных кислот.
Кетоновые тела образуются в печени и выполняют следующие функции:1. Энергетическая. Скелетная и сердечная мышцы, мозг и др внепеченочные ткани обеспечивают энергетические потребности за счет катаболизма кетоновых тел. Печень не может окислять кетоновые тела. 2.необходимы для образования миелиновых оболочек нервов и белого вещества головного мозга.
Утилизация кетоновых тел происходит в митохондриях. Бета-гидроксибутират превращается в ацетоацетат, а ацетоацетат вступает в реакцию с промежуточным продуктом ЦТК — сукцинил-КоА. При длительном голодании кетоновые тела становятся основным источником энергии для скелетных мышц, сердца и почек. Таким образом глюкоза сохраняется для окисления в мозге и эритроцитах. Уже через 2-3 дня после начала голодания концентрация кетоновых тел в крови достаточна для того, чтобы они проходили в клетки мозга и окислялись, снижая его потребности в глюкозе. β-Гидроксибутират (рис. 8-34), попадая в клетки, дегидрируется NAD-зависимой дегидрогеназой и превращается в ацетоацетат. Ацетоацетат активируется, взаимодействуя с сук-цинил-КоА — донором КоА: Ацетоацетат + Сукцинил-КоА → Ацетоацетил- КоА + Сукцинат
Реакцию катализирует сукцинил-КоА-ацето-ацетат-КоА-трансфераза. Этот фермент не синтезируется в печени, поэтому печень не использует кетоновые тела как источники энергии, а производит их «на экспорт». Кетоновые тела — хорошие топливные молекулы; окисление одной молекулы β-гидроксибутирата до СО2 и Н2О обеспечивает синтез 27 молекул АТФ. Эквивалент одной макроэргической связи АТФ (в молекуле сукцинил-КоА) используется на активацию ацетоацетата, поэтому суммарный выход АТФ при окислении одной молекулы β-гидроксибутирата — 26 молекул.
В норме процессы синтеза и использования кетоновых тел уравновешены, поэтому концентрация кетоновых тел в крови и в тканях обычно очень низка, и составляет 0,12-0,30 ммоль/л.. В норме в крови кол-во КТ 1-3 мг, в моче 40мг. При сахарном диабете 10-50 мг в моче. Накопление кетоновых тел в организме называется кетозом.Он сопровождается кетонемией и кетонурией. Физиологиеский кетоз при голодании, тяжелой мышечной работе, у новорожденных. Патологический при сахарном диабете. Однако при общем или при углеводном голодании может нарушаться баланс между образованием и утилизацией кетоновых тел. Это связано с тем, что скорость образования кетоновых тел зависит от скорости -окисления жирных кислот в печени, а процесс -окисления ускоряется при усилении липолиза (распада жира) в жировой ткани. Усиление липолиза может происходить под действием гормона адреналина, при мышечной работе, при голодании. При недостатке инсулина (сахарный диабет) также происходит усиление липолиза. При усилении липолиза увеличивается скорость утилизации кетоновых тел, которые являются важными источниками энергии при мышечной работе, голодании.
Постепенное истощение запасов углеводов при сахарном диабете приводит к относительному отставанию утилизации кетоновых тел от кетогенеза. Причина отставания: не хватает сукцинил-КоА и ЩУК, которые, в основном, являются продуктом обмена углеводов. Поэтому верно выражение: «Жиры сгорают в пламени углеводов». Это означает, что для эффективного использования продуктов распада жира необходимы продукты углеводного обмена: сукцинил-КоА и ЩУК.
Т.о., при углеводном голодании концентрация кетоновых тел в крови увеличивается. На 3-й день голодания концентрация кетоновых тел в крови будет примерно 2-3 ммоль/л, а при дальнейшем голодании — гораздо более высокой. Это состояние называют гиперкетонемия. У здоровых людей при мышечной работе и при голодании наблюдается гиперкетонемия, но она незначительна.
Похожая ситуация характерна для сахарного диабета. При сахарном диабете клетки постоянное сильнейшее углеводное голодание, потому что глюкоза плохо проникает в клетки. Наблюдается активация липолиза и повышается образование кетоновых тел. При тяжелых формах сахарного диабета концентрация кетоновых тел в крови может быть еще выше, и достигать опасных для жизни значений: до 20 ммоль/л и более. Все кетоновые тела являются органическими кислотами. Их накопление приводит к сдвигу pH в кислую сторону. В клинике повышение концентрации кетоновых тел в крови называется гиперкетонемия, а сдвиг pH при этом в кислую сторону — кетоацидоз. Нарушается работа многих ферментативных систем. Увеличение концентрации ацетоацетата приводит к ускоренному образованию ацетона. Ацетон — токсичное вещество (органический растворитель). Он растворяется в липидных компонентах клеточных мембран и дезорганизует их. Страдают все ткани организма, а больше всего — клетки нервной ткани. Это может проявляться потерей сознания (гипергликемическая кома). В очень тяжелых случаях может наступить гибель организма. Организм пытается защититься, поэтому часть кетоновых тел удаляется с мочой. Появление кетоновых тел в моче — это кетонурия. Для распознавания гипер- и гипогликемической комы применяется экспресс-диагностика кетоновых тел. Основана на том, что гиперкетонемия приводит к выведению кетоновых тел с мочой (кетонурия). Поэтому проводят цветную реакцию на наличие кетоновых тел в моче. Раньше диагноз ставили по запаху ацетона изо рта больного при гипергликемической коме (запах гнилых яблок).
источник
Тема: «БИОХИМИЯ ПЕЧЕНИ»
1. Химический состав печени: содержание гликогена, липидов, белков, минеральный состав.
2. Роль печени в углеводном обмене: поддержание постоянной концентрации глюкозы, синтез и мобилизация гликогена, глюконеогенез, основные пути превращения глюкозо-6-фосфата, взаимопревращения моносахаридов.
3. Роль печени в обмене липидов: синтез высших жирных кислот, ацилглицеролов, фосфолипидов, холестерола, кетоновых тел, синтез и обмен липопротеинов, понятие о липотропном эффекте и липотропных факторах.
4. Роль печени в белковом обмене: синтез специфических белков плазмы крови, образование мочевины и мочевой кислоты, холина, креатина, взаимопревращения кетокислот и аминокислот.
5. Метаболизм алкоголя в печени, жировое перерождение печени при злоупотреблении алкоголем.
6. Обезвреживающая функция печени: стадии (фазы) обезвреживания токсических веществ в печени.
7. Обмен билирубина в печени. Изменения содержания желчных пигментов в крови, моче и кале при различных видах желтух (надпечёночной, паренхиматозной, обтурационной).
8. Химический состав желчи и её роль; факторы, способствующие образованию желчных камней.
31.1. Функции печени.
Печень является органом, занимающим уникальное место в обмене веществ. В каждой печёночной клетке содержится несколько тысяч ферментов, катализирующих реакции многочисленных метаболических путей. Поэтому печень выполняет в организме целый ряд метаболических функций. Важнейшими из них являются:
- биосинтез веществ, которые функционируют или используются в других органах. К этим веществам относятся белки плазмы крови, глюкоза, липиды, кетоновые тела и многие другие соединения;
- биосинтез конечного продукта азотистого обмена в организме — мочевины;
- участие в процессах пищеварения — синтез желчных кислот, образование и экскреция желчи;
- биотрансформация (модификация и конъюгация) эндогенных метаболитов, лекарственных препаратов и ядов;
- выделение некоторых продуктов метаболизма (желчные пигменты, избыток холестерола, продукты обезвреживания).
31.2. Роль печени в обмене углеводов.
Основная роль печени в обмене углеводов заключается в поддержании постоянного уровня глюкозы в крови. Это осуществляется путём регуляции соотношения процессов образования и утилизации глюкозы в печени.
В клетках печени содержится фермент глюкокиназа, катализирующий реакцию фосфорилирования глюкозы с образованием глюкозо-6-фосфата. Глюкозо-6-фосфат является ключевым метаболитом углеводного обмена; основные пути его превращения представлены на рисунке 1.
31.2.1. Пути утилизации глюкозы. После приёма пищи большое количество глюкозы поступает в печень по воротной вене. Эта глюкоза используется прежде всего для синтеза гликогена (схема реакций приводится на рисунке 2). Содержание гликогена в печени здоровых людей обычно составляет от 2 до 8% массы этого органа.
Гликолиз и пентозофосфатный путь окисления глюкозы в печени служат в первую очередь поставщиками метаболитов-предшественников для биосинтеза аминокислот, жирных кислот, глицерола и нуклеотидов. В меньшей степени окислительные пути превращения глюкозы в печени являются источниками энергии для обеспечения биосинтетических процессов.
Рисунок 1. Главные пути превращения глюкозо-6-фосфата в печени. Цифрами обозначены: 1 — фосфорилирование глюкозы; 2 — гидролиз глюкозо-6-фосфата; 3 — синтез гликогена; 4 — мобилизация гликогена; 5 — пентозофосфатный путь; 6 — гликолиз; 7 — глюконеогенез.
Рисунок 2. Схема реакций синтеза гликогена в печени.
Рисунок 3. Схема реакций мобилизации гликогена в печени.
31.2.2. Пути образования глюкозы. В некоторых условиях (при голодании низкоуглеводной диете, длительной физической нагрузке) потребность организма в углеводах превышает то количество, которое всасывается из желудочно-кишечного тракта. В таком случае образование глюкозы осуществляется с помощью глюкозо-6-фосфатазы, катализирующей гидролиз глюкозо-6-фосфата в клетках печени. Непосредственным источником глюкозо-6-фосфата служит гликоген. Схема мобилизации гликогена представлена на рисунке 3.
Мобилизация гликогена обеспечивает потребности организма человека в глюкозе на протяжении первых 12 — 24 часов голодания. В более поздние сроки основным источником глюкозы становится глюконеогенез — биосинтез из неуглеводных источников.
Основными субстратами для глюконеогенеза служат лактат, глицерол и аминокислоты (за исключением лейцина). Эти соединения сначала превращаются в пируват или оксалоацетат — ключевые метаболиты глюконеогенеза.
Глюконеогенез — процесс, обратный гликолизу. При этом барьеры, создаваемые необратимыми реакциями гликолиза, преодолеваются при помощи специальных ферментов, катализирующих обходные реакции (см. рисунок 4).
Из других путей обмена углеводов в печени следует отметить превращение в глюкозу других пищевых моносахаридов — фруктозы и галактозы.
Рисунок 4. Гликолиз и глюконеогенез в печени.
Ферменты, катализирующие необратимые реакции гликолиза: 1 — глюкокиназа; 2 — фосфофруктокиназа; 3 — пируваткиназа.
Ферменты, катализирующие обходные реакции глюконеогенеза: 4 -пируваткарбоксилаза; 5 — фосфоенолпируваткарбоксикиназа; 6 -фруктозо-1,6-дифосфатаза; 7 — глюкозо-6-фосфатаза.
31.3. Роль печени в обмене липидов.
В гепатоцитах содержатся практически все ферменты, участвующие в метаболизме липидов. Поэтому паренхиматозные клетки печени в значительной степени контролируют соотношение между потреблением и синтезом липидов в организме. Катаболизм липидов в клетках печени протекает главным образом в митохондриях и лизосомах, биосинтез — в цитозоле и эндоплазматическом ретикулуме. Ключевым метаболитом липидного обмена в печени является ацетил-КоА, главные пути образования и использования которого показаны на рисунке 5.
Рисунок 5. Образование и использование ацетил-КоА в печени.
31.3.1. Метаболизм жирных кислот в печени. Пищевые жиры в виде хиломикронов поступают в печень через систему печёночной артерии. Под действием липопротеинлипазы, находящейся в эндотелии капилляров, они расщепляются до жирных кислот и глицерола. Жирные кислоты, проникающие в гепатоциты, могут подвергаться окислению, модификации (укорочению или удлинению углеродной цепи, образованию двойных связей) и использоваться для синтеза эндогенных триацилглицеролов и фосфолипидов.
31.3.2. Синтез кетоновых тел. При β-окислении жирных кислот в митохондриях печени образуется ацетил-КоА, подвергающийся дальнейшему окислению в цикле Кребса. Если в клетках печени имеется дефицит оксалоацетата (например, при голодании, сахарном диабете), то происходит конденсация ацетильных групп с образованием кетоновых тел (ацетоацетат,β-гидроксибутират, ацетон). Эти вещества могут служить энергетическими субстратами в других тканях организма (скелетные мышцы, миокард, почки, при длительном голодании — головной мозг). Печень не утилизирует кетоновые тела. При избытке кетоновых тел в крови развивается метаболический ацидоз. Схема образования кетоновых тел — на рисунке 6.
Рисунок 6. Синтез кетоновых тел в митохондриях печени.
31.3.3. Образование и пути использования фосфатидной кислоты. Общим предшественником триацилглицеролов и фосфолипидов в печени является фосфатидная кислота. Она синтезируется из глицерол-3-фосфата и двух ацил-КоА — активных форм жирных кислот (рисунок 7). Глицерол-3-фосфат может образоваться либо из диоксиацетонфосфата (метаболит гликолиза), либо из свободного глицерола (продукт липолиза).
Рисунок 7. Образование фосфатидной кислоты (схема).
Для синтеза фосфолипидов (фосфатидилхолина) из фосфатидной кислоты необходимо поступление с пищей достаточного количества липотропных факторов (веществ, препятствующих развитию жировой дистрофии печени). К этим факторам относятся холин, метионин, витамин В 12 , фолиевая кислота и некоторые другие вещества. Фосфолипиды включаются в состав липопротеиновых комплексов и принимают участие в транспорте липидов, синтезированных в гепатоцитах, в другие ткани и органы. Недостаток липотропных факторов (при злоупотреблении жирной пищей, хроническом алкоголизме, сахарном диабете) способствует тому, что фосфатидная кислота используется для синтеза триацилглицеролов (нерастворимых в воде). Нарушение образования липопротеинов приводит к тому, что избыток ТАГ накапливается в клетках печени (жировая дистрофия) и функция этого органа нарушается. Пути использования фосфатидной кислоты в гепатоцитах и роль липотропных факторов показаны на рисунке 8.
Рисунок 8. Использование фосфатидной кислоты для синтеза триацилглицеролов и фосфолипидов. Липотропные факторы обозначены знаком * .
31.3.4. Образование холестерола. Печень является основным местом синтеза эндогенного холестерола. Это соединение необходимо для построения клеточных мембран, является предшественником желчных кислот, стероидных гормонов, витамина Д 3 . Первые две реакции синтеза холестерола напоминают синтез кетоновых тел, но протекают в цитоплазме гепатоцита. Ключевой фермент синтеза холестерола — β-гидрокси-β-метилглутарил-КоА-редуктаза (ГМГ-КоА-редуктаза)ингибируется избытком холестерола и желчными кислотами по принципу отрицательной обратной связи (рисунок 9).
Рисунок 9. Синтез холестерола в печени и его регуляция.
31.3.5. Образование липопротеинов. Липопротеины — белково-липидные комплексы, в состав которых входят фосфолипиды, триацилглицеролы, холестерол и его эфиры, а также белки (апопротеины). Липопротеины транспортируют нерастворимые в воде липиды к тканям. В гепатоцитах образуются два класса липопротеинов — липопротеины высокой плотности (ЛПВП) и липопротеины очень низкой плотности (ЛПОНП).
31.4. Роль печени в обмене белков.
Печень является органом, регулирующим поступление азотистых веществ в организм и их выведение. В периферических тканях постоянно протекают реакции биосинтеза с использованием свободных аминокислот, либо выделение их в кровь при распаде тканевых белков. Несмотря на это, уровень белков и свободных аминокислот в плазме крови остаётся постоянным. Это происходит благодаря тому, что в клетках печени имеется уникальный набор ферментов, катализирующих специфические реакции обмена белков.
31.4.1. Пути использования аминокислот в печени. После приёма белковой пищи в клетки печени по воротной вене поступает большое количество аминокислот. Эти соединения могут претерпевать в печени ряд превращений, прежде чем поступить в общий кровоток. К этим реакциям относятся (рисунок 10):
а) использование аминокислот для синтеза белков;
б) трансаминирование — путь синтеза заменимых аминокислот; осуществляет также взаимосвязь обмена аминокислот с глюконеогенезом и общим путём катаболизма;
в) дезаминирование — образование α-кетокислот и аммиака;
г) синтез мочевины — путь обезвреживания аммиака (схему см. в разделе «Обмен белков»);
д) синтез небелковых азотсодержащих веществ (холина, креатина, никотинамида, нуклеотидов и т.д.).
Рисунок 10. Обмен аминокислот в печени (схема).
31.4.2. Биосинтез белков. В клетках печени синтезируются многие белки плазмы крови: альбумины (около 12 г в сутки), большинство α- и β-глобулинов, в том числе транспортные белки (ферритин, церулоплазмин, транскортин, ретинолсвязывающий белок и др.). Многие факторы свёртывания крови (фибриноген, протромбин, проконвертин, проакцелерин и др.) также синтезируются в печени.
31.5. Обезвреживающая функция печени.
В печени обезвреживаются неполярные соединения различного происхождения, в том числе эндогенные вещества, лекарственные препараты и яды. Процесс обезвреживания веществ включает две стадии (фазы):
1) фаза модификации — включает реакции окисления, восстановления, гидролиза; для ряда соединений необязательна;
2) фаза конъюгации — включает реакции взаимодействия веществ с глюкуроновои и серной кислотами, глицином, глутаматом, таурином и другими соединениями.
Более подробно реакции обезвреживания будут рассмотрены в разделе «Биотрансформация ксенобиотиков».
31.6. Желчеобразовательная функция печени.
Желчь — жидкий секрет желтовато-коричневого цвета, выделяемый печёночными клетками (500-700 мл в сутки). В состав желчи входят: желчные кислоты, холестерол и его эфиры, желчные пигменты, фосфолипиды, белки, минеральные вещества (Nа + , К + , Са 2+ , Сl — ) и вода.
31.6.1. Желчные кислоты. Являются продуктами метаболизма холестерола, образуются в гепатоцитах. Различают первичные (холевая, хенодезоксихолевая) и вторичные (дезоксихолевая, литохолевая) желчные кислоты. В желчи присутствуют главным образом желчные кислоты, конъюгированные с глицином или таурином (например, гликохолевая, кислота, таурохолевая кислота и т.д.).
Желчные кислоты принимают непосредственное участие в переваривании жиров в кишечнике:
- оказывают на пищевые жиры эмульгирующее действие;
- активируют панкреатическую липазу;
- способствуют всасыванию жирных кислот и жирорастворимых витаминов;
- стимулируют перистальтику кишечника.
При нарушении оттока желчи желчные кислоты проникают в кровь и мочу.
31.6.2. Холестерол. С желчью выводится из организма избыток холестерола. Холестерол и его эфиры присутствуют в желчи в виде комплексов с желчными кислотами (холеиновые комплексы). При этом отношение содержания желчных кислот к содержанию холестерола (холатный коэффициент) должно быть не ниже 15. В противном случае нерастворимый в воде холестерол выпадает в осадок и откладывается в виде камней желчного пузыря (желчно-каменная болезнь).
31.6.3. Желчные пигменты. Из пигментов в желчи преобладает конъюгированный билирубин (моно- и диглюкуронид билирубина). Он образуется в клетках печени в результате взаимодействия свободного билирубина с УДФ-глюкуроновой кислотой. При этом снижается токсичность билирубина и увеличивается его растворимость в воде; далее конъюгированный билирубин секретируется в желчь. При нарушении оттока желчи (механическая желтуха) в крови значительно увеличивается содержание прямого билирубина, в моче обнаруживается билирубин, в кале и моче снижено содержание стеркобилина. Дифференциальную диагностику желтух см. в разделе «Обмен сложных белков».
31.6.4. Ферменты. Из ферментов, обнаруженных в желчи, следует в первую очередь отметить щелочную фосфатазу. Это экскреторный фермент, синтезируемый в печени. При нарушении оттока желчи активность щелочной фосфатазы в крови возрастает.
источник
В гепатоцитах содержатся практически все ферменты, участвующие в метаболизме липидов. Поэтому паренхиматозные клетки печени в значительной степени контролируют соотношение между потреблением и синтезом липидов в организме. Катаболизм липидов в клетках печени протекает главным образом в митохондриях и лизосомах, биосинтез — в цитозоле и эндоплазматическом ретикулуме. Ключевым метаболитом липидного обмена в печени является ацетил-КоА, главные пути образования и использования которого показаны на рисунке 5.
Рисунок 5. Образование и использование ацетил-КоА в печени.
Метаболизм жирных кислот в печени. Пищевые жиры в виде хиломикронов поступают в печень через систему печёночной артерии. Под действиемлипопротеинлипазы, находящейся в эндотелии капилляров, они расщепляются до жирных кислот и глицерола. Жирные кислоты, проникающие в гепатоциты, могут подвергаться окислению, модификации (укорочению или удлинению углеродной цепи, образованию двойных связей) и использоваться для синтеза эндогенных триацилглицеролов и фосфолипидов.
Синтез кетоновых тел. При β-окислении жирных кислот в митохондриях печени образуется ацетил-КоА, подвергающийся дальнейшему окислению в цикле Кребса. Если в клетках печени имеется дефицит оксалоацетата (например, при голодании, сахарном диабете), то происходит конденсация ацетильных групп с образованием кетоновых тел (ацетоацетат,β-гидроксибутират, ацетон). Эти вещества могут служить энергетическими субстратами в других тканях организма (скелетные мышцы, миокард, почки, при длительном голодании — головной мозг). Печень не утилизирует кетоновые тела. При избытке кетоновых тел в крови развивается метаболический ацидоз. Схема образования кетоновых тел — на рисунке 6.
Рисунок 6. Синтез кетоновых тел в митохондриях печени.
Образование и пути использования фосфатидной кислоты. Общим предшественником триацилглицеролов и фосфолипидов в печени является фосфатидная кислота. Она синтезируется из глицерол-3-фосфата и двух ацил-КоА — активных форм жирных кислот (рисунок 7). Глицерол-3-фосфат может образоваться либо из диоксиацетонфосфата (метаболит гликолиза), либо из свободного глицерола (продукт липолиза).
Рисунок 7. Образование фосфатидной кислоты (схема).
Для синтеза фосфолипидов (фосфатидилхолина) из фосфатидной кислоты необходимо поступление с пищей достаточного количества липотропных факторов (веществ, препятствующих развитию жировой дистрофии печени). К этим факторам относятся холин, метионин, витамин В12, фолиевая кислота и некоторые другие вещества. Фосфолипиды включаются в состав липопротеиновых комплексов и принимают участие в транспорте липидов, синтезированных в гепатоцитах, в другие ткани и органы. Недостаток липотропных факторов (при злоупотреблении жирной пищей, хроническом алкоголизме, сахарном диабете) способствует тому, что фосфатидная кислота используется для синтеза триацилглицеролов (нерастворимых в воде). Нарушение образования липопротеинов приводит к тому, что избыток ТАГ накапливается в клетках печени (жировая дистрофия) и функция этого органа нарушается. Пути использования фосфатидной кислоты в гепатоцитах и роль липотропных факторов показаны на рисунке 8.
Рисунок 8. Использование фосфатидной кислоты для синтеза триацилглицеролов и фосфолипидов. Липотропные факторы обозначены знаком * .
Образование холестерола. Печень является основным местом синтеза эндогенного холестерола. Это соединение необходимо для построения клеточных мембран, является предшественником желчных кислот, стероидных гормонов, витамина Д3. Первые две реакции синтеза холестерола напоминают синтез кетоновых тел, но протекают в цитоплазме гепатоцита. Ключевой фермент синтеза холестерола — β-гидрокси-β-метилглутарил-КоА-редуктаза (ГМГ-КоА-редуктаза) ингибируется избытком холестерола и желчными кислотами по принципу отрицательной обратной связи (рисунок 9).
Рисунок 9. Синтез холестерола в печени и его регуляция.
Образование липопротеинов. Липопротеины — белково-липидные комплексы, в состав которых входят фосфолипиды, триацилглицеролы, холестерол и его эфиры, а также белки (апопротеины). Липопротеины транспортируют нерастворимые в воде липиды к тканям. В гепатоцитах образуются два класса липопротеинов — липопротеины высокой плотности (ЛПВП) и липопротеины очень низкой плотности (ЛПОНП).
источник
10. Пути обмена Ацетил-КоА, значение каждого пути. Общая характеристика процесса биосинтез жирных кислот. Понятие об эссенциальных жирных кислотах и их роли в профилактике нарушений обмена липидов.
Ацетил-КоА образуется в специфических реакциях катаболизма жирных кислот и некоторых аминокислот. В реакциях гликолиза из глюкозы образуется ПВК, которая поступает в матрикс митохондрий и превращается в Ацетил-КоА с участием ПВК ДГ. Так как внутренняя мембрана митохондрий непроницаема для Ацетил-КоА, поэтому он при участии цитратсинтазы конденсируется с ЩУК с образованием цитрата: [Ацетил-КоА + Оксалоацетат → Цитрат + HS-КоА]. Затем транслоказа переносит цитрат в цитоплазму. Перенос цитрата в цитоплазму происходит только при увеличении количества цитрата в митохондриях, когда изоцитратдегидрогеназа и α-кетоглутаратдегидрогеназа ингибированы высокими концентрациями НАДН2 и АТФ (при избытке углеводов и низком энергопотреблении). В цитоплазме цитрат расщепляется под действием фермента цитрат-лиазы: [Цитрат + HSKoA + АТФ → Ацетил-КоА + АДФ+ Pн + ЩУК].
Биосинтез жирных кислот протекает в печени с участием НАДФН, АТФ, Мn 2+ и НСО3– (в качестве источника СО2), витаминов (биотин, РР); субстратом является ацетил-КоА, конечным продуктом – пальмитиновая кислота.
Эссенциальные жирные кислоты (Омега-3,6) — комплекс полиненасыщенных жирных кислот, которые принимают значительное участие в метаболизме человека. К ним относят линолевую и линоленовую к-ты. Жирные кислоты омега-6 понижают уровень холестерина в крови, а жирные кислоты омега-3 — уровень триглицерида и кровяное давление.
11. Кетоновые тела: биологическая роль, реакции обмена, регуляция. Кетонемия, кетонурия, причины и механизмы развития, последствия.
При дефиците углеводов наблюдается: ↓ие распада глюкозы; содержание ПВК ↓ся; содиржание ЩУК ↓ся; ЦТК угнетается. ↑ся распад ЖК с образованием ацетил-S-KoA. Роль кетоновых тел резко ↑ся при голодании – они служат источником энергии для мышц и мозга при голодании. Окисление кетоновых тел в тканях: [β-гидроксибутират (β-гидроксибутират-дегидрогеназа)↔ ацетоацетат (Сукцинил-КоА-ацетоацетат-КоА-трансфераза)→ Ацетоацетил-КоА (Тиолаза)↔ 2-ацетил-КоА→ ЦТК]. В норме кетоновых тел ни в крови ни в моче нет. Кетонемию и кетонурию наблюдают при СД, углеводном голодании, лихорадочных состояниях, общем голодании и истощении, тяжелой интоксикации свинцом. Следствием кетонемии является кетоацидоз и ацетоновое отравление (ацетон растворяет структурные липиды клеток), при котором нарушается транспорт глюкозы через биомембраны и резко угнетается деятельность ЦНС.
12. Функции холестерина. Фонд холестерина организма: пути поступления, использования и выведения. Синтез холестерина: основные этапы, регуляция процесса.
Холестерол — стероид, характерный только для животных организмов. Он синтезируется во многих тканях человека, но основное место синтеза — печень. Холестерол выполняет много функций: входит в состав всех мембран клеток и влияет на их свойства, служит исходным субстратом в синтезе жёлчных кислот и стероидных гормонов. Ежедневно из организма выводится около 1 г холестерола. Приблизительно половина этого количества экскретируется с фекалиями после превращения в желчные кислоты. Оставшаяся часть выводится в виде нейтральных стероидов. Большая часть холестерола, поступившего в желчь, реабсорбируется
Реакции синтеза холестерола происходят в цитозоле клеток. Сложный путь синтеза холестерола можно разделить на 3 этапа. Первый этап заканчивается образованием мевалоната. Две молекулы ацетил-КоА конденсируются ферментом тиолазой с образованием ацетоацетил-КоА. Фермент гидроксиметилглутарил-КоА-синтаза присоединяет третий ацетильный остаток с образованием ГМГ-КоА (3-гидрокси-3-метилглутарил-КоА). Эта последовательность реакций сходна с начальными стадиями синтеза кетоновых тел. Однако реакции синтеза кетоновых тел происходят в митохондриях печени, а реакции синтеза холестерола — в цитозоле клеток. Следующая реакция, катализируемая ГМГ-КоА-редуктазой, является регуляторной в метаболическом пути синтеза холестерола. В этой реакции происходит восстановление ГМГ-КоА до мевалоната с использованием 2 молекул NADPH. Фермент ГМГ-КоА-редуктаза — гликопротеин, пронизывающий мембрану ЭР, активный центр которого выступает в цитозоль. На втором этапе синтеза мевалонат превращается в пятиуглеродную изопреноидную структуру, содержащую пирофосфат — изопентенилпирофосфат. Продукт конденсации 2 изопреновых единиц — геранилпирофосфат. Присоединение ещё 1 изопреновой единицы приводит к образованию фарнезилпирофосфата — соединения, состоящего из 15 углеродных атомов. Две молекулы фарнезилпирофосфата конденсируются с образованием сквалена — углеводорода линейной структуры, состоящего из 30 углеродных атомов. На третьем этапе синтеза холестерола сквален через стадию образования эпоксида ферментом циклазой превращается в молекулу ланостерола, содержащую 4 конденсированных цикла и 30 атомов углерода. Далее происходит 20 последовательных реакций, превращающих ланостерол в холестерол. На последних этапах синтеза от ланостерола отделяется 3 атома углерода, поэтому холестерол содержит 27 углеродных атомов.
источник
4.Пути использования ацетил КоА. Механизм образования и значение ацетоуксусной кислоты. Биосинтез кетоновых тел. Кетоацидоз.
На первой стадии из двух молекул ацетил-КоА образуется ацетоацетил-КоА:
Реакция катализируется ферментом ацетил-КоА-ацетилтрансферазой (ацетоацетил-КоА-тиолазой). Затем ацетоацетил-КоА взаимодействует еще с одной молекулой ацетил-КоА. Реакция протекает под влиянием фермента гидроксиметилглутарил-КоА-синтазы:
Образовавшийся β-гидрокси-β-метилглутарил-КоА способен под действием гидроксиметилглутарил-КоА-лиазы расщепляться на ацетоацетат и ацетил-КоА:
Ацетоуксусная кислота способна восстанавливаться при участии НАД-зависимой D-β-гидроксибутиратдегидрогеназы; при этом образуется D-β-гидроксимасляная кислота (D-β-гидроксибутират). Следует еще раз подчеркнуть, что фермент специфичен по отношению к D-стереоизомеру и не действует на КоА-эфиры:
Существует также и второй путь синтеза кетоновых тел. Образовавшийся путем конденсации двух молекул ацетил-КоА ацетоацетил-КоА способен отщеплять коэнзим А и превращаться в свободную ацетоуксусную кислоту. Этот процесс катализируется ферментом ацетоацетил-КоА-гидролазой (деацилазой):
Однако второй путь образования ацетоуксусной кислоты не имеет существенного значения, так как активность деацилазы в печени низкая. В крови здорового человека кетоновые тела содержатся лишь в очень небольших концентрациях. При патологических состояниях (например, у лиц с тяжелой формой сахарного диабета, при голодании, а также у животных с острым аллоксановым диабетом) концентрация кетоновых тел в крови увеличивается и может достигать 20 ммоль. Такое состояние, которое получило название кетоза, возникает в тех случаях, когда скорость образования кетоновых тел превышает способность периферических тканей их утилизировать.
В последние годы начали постепенно накапливаться данные, указывающие на важную роль кетоновых тел в поддержании энергетического гомеостаза. Кетоновые тела — своего рода поставщики топлива для мышц, мозга, почек и действуют, возможно, как часть регуляторного механизма с обратной связью, предотвращая чрезвычайную мобилизацию жирных кислот из жировых депо. Печень в этом смысле является исключением, она не использует кетоновые тела в качестве энергетического материала.
В периферических тканях β-гидроксимасляная кислота окисляется до ацетоуксусной кислоты, а последняя активируется с образованием соответствующего КоА-эфира (ацетоацетил-КоА).
Существует два ферментативных механизма активации ацетоуксусной кислоты. Первый путь -это использование АТФ и HS-КоА аналогично тому, как при активации жирных кислот:
Реакция катализируется ферментом ацил-КоА-синтетазой, или тиокиназой. Второй путь активации — это перенос КоА от сукцинил-КоА на ацетоуксусную кислоту:
Данная реакция катализируется ферментом сукцинил-КоА-ацетоацетат-трансферазой. Возможно, что биологически более важным является именно этот путь активации ацетоацетата.
Образовавшийся в ходе этих реакций ацетоацетил-КоА подвергается далее тиолитическому расщеплению с образованием двух молекул ацетил-КоА:
А уже ацетил-КоА, как это известно, окисляется в цикле трикарбоновых кислот (цикл Кребса) до СО2 и Н20.
Кетоацидоз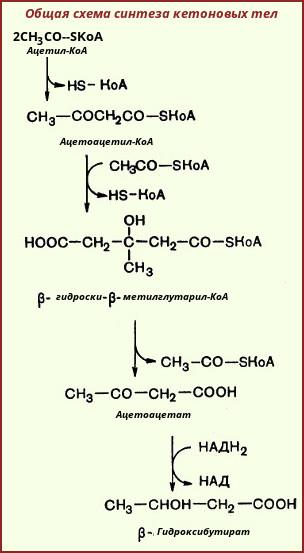
В норме концентрация КТ в крови составляет 1—3 мг/дл (до 0,2 мМоль/л), но при голодании значительно увеличивается. Увеличение концентрации КТ в крови называют кетонемией. При кетонемии развивается кетонурия — выделение КТ с мочой. Накопление КТ в организме приводит к кетоацидозу, так как КТ (кроме ацетона) являются водорастворимыми органическими кислотами (рК
Ацидоз достигает опасных величин при сахарном диабете, так как концентрация КТ при этом заболевании может доходить до 400—500 мг/дл. Тяжёлая форма ацидоза — одна из основных причин смерти при сахарном диабете.
Биосинтез триацилглицеринов: стадии процесса. Липотропные факторы и их роль в биосинтезе липидов в печени.
Биосинтез триацилглицеринов. Образующиеся в результате биосинтеза жирные кислоты в организмах животных и человека в свободном виде встре- чаются лишь в незначительных количествах, а присутствуют главным образом в виде триацилглицеринов. Синтез триацилглицеринов происходит в печени и жировой ткани из КоА-производных жирных кислот через фосфатидную ки- слоту по реакции:
2 R C SКоА + О Н2С ОН НС Н2С О РО3Н2 ОН Н2С О СH Н2С О РО3Н2 С R O R C O O + 2 HS-KoA Фосфатидная кислота Фосфорилирование глицерина осуществляется глицеролкиназой за счет энергии АТФ. Глицерол-3-фосфат может образовываться и при восстановлении диоксиацетонфосфата. Гидролиз фосфатидной кислоты фосфатазой приводит к образованию 1,2-диацилглицерина, который, реагируя с другой молекулой ацил-КоА, обра- зует нейтральный триацилглицерин. В слизистой кишечника триацилглицерины синтезируются из свободных кислот, моно- и диацилглицеринов, но эти процессы характерны только для слизистой оболочки кишечника. Перенос остатка жирной кислоты происходит через ацильное производное КоА.
Липотропные факторы – вещества, способствующие синтезу фосфолипидов и препятствующие отложению триацилглицеролов в тканях.
Липотропный эффект этих соединений связан с тем, что общим предшественником триацилглицеролов и фосфолипидов является фосфатидная кислота. При недостатке липотропных факторов фосфатидная кислота используется преимущественно для синтеза триацилглицеролов. Они нерастворимы в воде и накапливаются в клетках, способствуя их жировому перерождению. Запомните, какие липотропные факторы можно использовать в качестве препаратов для его предупреждения:
1) холин – самое распространённое азотистое основание фосфолипидов;
2) метионин – незаменимая аминокислота, предшественник S-аденозилметионина, универсального донора метильных групп;
3) фолиевая кислота и витамин В12 – предшественники коферментов, участвующих в реакциях переноса СН3-групп.
При введении липотропных факторов снижается синтез триацилглицеролов и усиливается выведение жира из клеток при участии липопротеиновых комплексов
источник
Жирные кислоты, поступающие в гепатоциты, активируются и подвергаются b-окислению с образованием ацетил-КоА. Именно этот ацетил-КоА используется для синтеза ацетоновых тел: ацетоацетата, b-гидроксибутирата и ацетона, по следующей схеме:
Образовавшиеся ацетоновые тела поступают из гепатоцитов в кровь и разносятся к клеткам различных органов. Этот процесс в той или иной мере идет постоянно и ацетоновые тела постоянно присутствуют в крови в концентрации до 30 мг/л. Ежесуточное их выделение с мочой не превышает 20 мг.
Ацетоновые тела в норме достаточно хорошо утилизируются клетками периферических тканей, в особенности это касается скелетных мышц и миокарда, которые значительную часть нужной им энергии получают за счет окисления ацетоновых тел. Лишь клетки центральной нервной системы в обычных условиях практически не утилизируют ацетоновые тела, однако при голодании даже головной мозг от 1/2 до 3/4 свой потребности в энергии может удовлетворять за счет окисления ацетоновых тел.
Ацетоацетат,поступающий в клетки различных тканей, прежде всего подвергается активации с помощью одного из двух возможных механизмов:
-поступающие в дальнейшем в цикл Кребса, где ацетильные остатки окисляются до СО2 и Н2О.
— Основным путем активации ацетоацетата в клетках является путь с участием тиафоразы. В гепатоцитах нет этого фермента. Именно поэтому образовавшийся в гепатоцитах ацетоацетат в них не активируется и не окисляется, тем самым создаются условия для » экспорта» ацетоацетата из гепатоцитов в кровь.
b-Гидроксибутират в клетках предварительно окисляется с участием НАД+ в ацетоацетат. Эта реакция катализируется ферментом b-гидроксибутиратдегидрогеназой.
Ацетон также может окисляться в клетках периферических органов. Возможны два варианта его окисления: во-первых, он может расщепляться до ацетильного и формильного остатков; во-вторых, через пропандиол он может превращаться в пируват.
Ацетоновые тела, накапливаясь в крови и в тканях, оказывают ингибирующее действие на липолиз, в особенности это касается расщепления триглицеридов в липоцитах. Биологическая роль этого регуляторного механизма становится понятной, если принять во внимание, что ацтоацетат и гидроксибутират представляют собой достаточно сильные органические кислоты, в связи с чем их избыточное накопление в крови приводит к развитию ацидоза. Снижение уровня липолиза в клетках жировой ткани приводит к уменьшению притока высших жирных кислот в гепатоциты и к снижению скорости образования ацетоновых тел и, следовательно, снижению их содержания в крови.
8 вопрос. Ацетил-КоА образуется в специфических реакциях катаболизма жирных кислот и некоторых аминокислот. Однако главным источником ацетил-КоА служит пировиноградная кислота, образующаяся в реакциях катаболизма глюкозы и некоторых аминокислот. Превращение пирувата в ацетил-КоА происходит при участии набора ферментов, структурно объединённых в пируватдегидрогеназный комплекс (ПДК). Ацетильный остаток — ацетил- Ко А далее окисляется в цикле лимонной кислоты до СО2 и Н2О. В этих реакциях окисления принимают участие NAD- и FAD-зависимые дегидрогеназы, поставляющие электроны и протоны в ЦПЭ, по которой они передаются на О2. это второй шаг кислородного клеточного дыхания, декарбоксилирования пирувата, который происходит в матриксе митохондрии. Ацетил-КоА затем поступает в цикл трикарбоновых кислот.
Ацетил-КоА является ключевым метаболитом липидного обмена в печени
Ацетил-КоА, являясь конечным продуктом гликолитического цикла, может использоваться как источник энергии (в цикле Кребса), а также участвовать в синтезе триглицеридов, холестерина, стероидов и образовании кетоновых тел
У животных ацетил-КоА является основой баланса между углеводным обменом и жировым обменом. Обычно ацетил-КоА из метаболизма жирных кислот поступает в цикл трикарбоновых кислот, содействуя энергетическому обеспечению клеток. В печени, когда уровень циркуляции жирных кислот высок, производство ацетил-КоА от разрыва жиров превышает энергетические потребности клетки. Чтобы использовать энергию, доступную из лишних ацетил-КоА, создаются кетоновые тела, которые затем могут циркулировать в крови. В некоторых обстоятельствах это может привести к высокому уровню кетоновых тел в крови, состоянию, называемому кетозом, которое отличается от кетоацидоза, опасного состояния, способного повлиять на диабетиков.
Синтез кетоновых тел. При β-окислении жирных кислот в митохондриях печени образуется ацетил-КоА, подвергающийся дальнейшему окислению в цикле Кребса. Если в клетках печени имеется дефицит оксалоацетата (например, при голодании, сахарном диабете), то происходит конденсация ацетильных групп с образованием кетоновых тел (ацетоацетат,β-гидроксибутират, ацетон). Эти вещества могут служить энергетическими субстратами в других тканях организма (скелетные мышцы, миокард, почки, при длительном голодании — головной мозг). Печень не утилизирует кетоновые тела. При избытке кетоновых тел в крови развивается метаболический ацидоз.
Две молекулы ацетил-КоА могут быть соединены, чтобы создать ацетоацетил-КоА, что будет первым шагом в ГМГ-КоА/биосинтезе холестерина, предшествующем синтезу изопреноидов. У животных ГМГ-КоА — это жизненный предшественник синтеза холестерина и кетоновых тел.
Ацетил-КоА может быть карбоксилирован в цитозоле в ацетил-КоА карбоксилазу, давая начало малонил-КоА, необходимого для синтеза флавоноидов и родственных поликетидов, для удлинения жирных кислот (образование восков),
У животных организмов ацетил КоА играет роль в обмене веществ, по мимо этого КоА как баланс между жировым обменом, так и углеводным. Для содействия клеток энергетическому соединению, КоА из жирных кислот поступает в цикл трикорбоновых кислот.
Субстрат для синтеза жирных кислот
Ацетил-КоА, образующийся под действием пируватдегидрогеназного комплекса (1), не проникает через мембрану митохондрий в цитоплазму, где происходит синтез жирных кислот. Поэтому ацетил-КоА конденсируется ферментом цитратсинтазой с оксалоацетатом (2), образуя цитрат, который с помощью транслоказы переносится в цитоплазму, где он расщепляется под действием цитратлиазы (3), и ацетил-КоА используется как субстрат для синтеза жирных кислот.
Первая реакция синтеза жирных кислот — это превращение ацетил-КоА в малонил-КоА:
 Фермент, катализирующий эту реакцию, ацетил-КоА-карбоксилаза,является регуляторным в биосинтезе жирных кислот. Он относится к классу лигаз и в качестве кофермента использует биотин.
Фермент, катализирующий эту реакцию, ацетил-КоА-карбоксилаза,является регуляторным в биосинтезе жирных кислот. Он относится к классу лигаз и в качестве кофермента использует биотин.
Исходным субстратом для синтеза холестерола является ацетил-КоА.Синтез холестерола происходит в абсорбтивный период, когда в печени активируется гликолиз, который является главным источником ацетил-КоА для синтеза холестерола. В синтезе холестерола из ацетил-КоА участвует около 30 ферментов, и этот метаболический путь является одним из самых длинных в организме. Реакции синтеза холестерола происходят в цитозоле и эндоплазматическом ретикулуме. Для синтеза необходим ацетил-КоА и NАDPН, образующийся в основном в реакциях пентозофосфатного пути катаболизма глюкозы. Все 27 атомов углерода холестерола происходят из ацетил-КоА.
2. Процесс синтеза холестерола условно разделяют на три этапа. I этап — реакции от ацетил-КоА до мевалоновой кислоты.
Так как ацетил-КоА образуется в митохондриях, а синтез холестерола происходит в цитоплазме клеток, то (как и при синтезе жирных кислот) ацетилКоА доставляется в цитоплазму в составе цитрата. Первые реакции процесса сходны с реакциями синтеза кетоновых тел, которые, в отличие от синтеза холестерола, протекают в митохондриях.


9 вопрос. Биосинтез жирных кислот наиболее активно происходит в цитозоле клеток печени, кишечника, жировой ткани в состоянии покоя или после еды.
Условно можно выделить 4 этапа биосинтеза:
1. Образование ацетил-SКоА из глюкозы, других моносахаров или кетогенных аминокислот.
2. Перенос ацетил-SКоА из митохондрий в цитозоль:*может быть в комплексе с карнитином, подобно тому как переносятся внутрь митохондрии высшие жирные кислоты, но здесь транспорт идет в другом направлении, *обычно в составе лимонной кислоты, образующейся в первой реакции ЦТК.
П
 оступающий из митохондрий цитрат в цитозоле расщепляется АТФ-цитрат-лиазой до оксалоацетата и ацетил-SКоА. Оксалоацетат в дальнейшем восстанавливается до малата, и последний либо переходит в митохондрии (малат-аспартатный челнок), либо декарбоксилируется в пируватмалик-ферментом («яблочный» фермент).
оступающий из митохондрий цитрат в цитозоле расщепляется АТФ-цитрат-лиазой до оксалоацетата и ацетил-SКоА. Оксалоацетат в дальнейшем восстанавливается до малата, и последний либо переходит в митохондрии (малат-аспартатный челнок), либо декарбоксилируется в пируватмалик-ферментом («яблочный» фермент).
3. Образование малонил-SКоА из ацетил-SКоА. Карбоксилирование ацетил-SКоА катализируется ацетил-SКоА-карбоксилазой, мульферментным комплексом из трех ферментов.
4. Синтез пальмитиновой кислоты. Осуществляется мультиферментным комплексом «синтаза жирных кислот» (синоним пальмитатсинтаза) в состав которого входит 6 ферментов и ацил-переносящий белок (АПБ). Ацил-переносящий белок включает производное пантотеновой кислоты – 6-фосфопантетеин (ФП), имеющий HS-группу, подобно HS-КоА. Один их ферментов комплекса, 3-кетоацил-синтаза, также имеет HS-группу в составе цистеина. Взаимодействие этих групп обусловливает начало и продолжение биосинтеза жирной кислоты, а именно пальмитиновой кислоты. Для реакций синтеза необходим НАДФН. В первых двух реакциях последовательно присоединяются малонил-SКоА к фосфопантетеину ацил-переносящего белка и ацетил-SКоА к цистеину 3-кетоацилсинтазы. 3-Кетоацилсинтаза катализирует третью реакцию – перенос ацетильной группы на С2 малонила с отщеплением карбоксильной группы. Далее кетогруппа в реакциях восстановления (3-кетоацил-редуктаза), дегидратации (дегидратаза) и опять восстановления (еноил-редуктаза) превращается в метиленовую с образованием насыщенного ацила, связанного с фосфопантетеином. Ацилтрансфераза переносит полученный ацил на цистеин 3-кетоацил-синтазы, к фосфопантетеину присоединяется малонил-SКоА и цикл повторяется 7 раз до образования остатка пальмитиновой кислоты. После этого пальмитиновая кислота отщепляется шестым ферментом комплекса тиоэстеразой.
Удлинение цепи жирных кислот
С интезированная пальмитиновая кислота при необходимости поступает в эндоплазматический ретикулум или в митохондрии. Здесь с участием малонил-S-КоА и НАДФН цепь удлиняется до С18 или С20. Удлиняться могут и ненасыщенные жирные кислоты (олеиновая, линолевая, линоленовая) с образованием производных эйкозановой кислоты (С20). Но двойная связь животными клетками вводится не далее 9 атома углерода, поэтому ω3- и ω6-полиненасыщенные жирные кислоты синтезируются только из соответствующих предшественников.
интезированная пальмитиновая кислота при необходимости поступает в эндоплазматический ретикулум или в митохондрии. Здесь с участием малонил-S-КоА и НАДФН цепь удлиняется до С18 или С20. Удлиняться могут и ненасыщенные жирные кислоты (олеиновая, линолевая, линоленовая) с образованием производных эйкозановой кислоты (С20). Но двойная связь животными клетками вводится не далее 9 атома углерода, поэтому ω3- и ω6-полиненасыщенные жирные кислоты синтезируются только из соответствующих предшественников.
Синтезир-ся в орг-ме в основном пальмитиновая к-та. При необходимости ж к-ты с большим числом углеродных атомов. Ненасыщенные ж к-ты обр-ся на мембранах ЭПС с участием О2, НАДН и В5. Под воздействием ферментов десатураз обр-ся пальмитиновая и олеиновая к-ты. Полиненасыщенные ж к-ты (линолевая, арахидоновая, линоленовая) должны поступать с пищей. Источником углевода для синтеза ж к-т служит ацетилКоА, обр-ся при распаде углеводов. Избыток углеводов, поступающих в орг-м трансформир-ся в ж к-ты, а затем в жиры.
Лимитирующим ферментом является ацетил-КоАкарбоксилаза. Аллостерические активаторы — АТФ и цитрат, ингибиторы — жирные кислоты с длинной цепью. Инсулин, эстрогены активиру¬ют, катехоламины и стресс ингибируют синтез жирных кислот. Значение:при распаде УВ обр ацетил-Коа, который используется в синтезе ЖК, т.о. избыток УВ запасается в виде жира.
источник