Составитель текста – Анисимова Е.С..
Авторские права защищены (продавать текст нельзя). Курсив не зубрить.
Замечания можно присылать по почте: exam_bch@mail.ru
https://vk.com/bch_5
ПАРАГРАФ 89:
«Белки и пептиды плазмы крови»
О функциях глобулинов см. также п.39. О пептидах см. п. 56.
Содержание параграфа:
Пептиды плазмы.
Гемоглобин
Функциональные и нефункциональные белки плазмы.
Фракции белков плазмы крови.
Отклонения содержания белков плазмы от нормы.
Дис/протеин/емии.
Гипо/альбумин/емии.
Ф У Н К Ц И И белков плазмы.
К пептидам, которые есть в крови в норме, относятся
ангиотензин (8 аминоацилов) и ряд других белково-пептидных гормонов, в составе которых менее 100 аминоацилов:
инсулин (51 аминоацил), глюкагон (29 аминоацилов) и т.д.. Но свойствам инсулин является типичным белком.
Гемоглобин
не относится к белкам плазмы крови (БПК) –
в норме он находится внутри эритроцитов (п.121),
а в плазму попадает при их разрушении (при гемолизе);
выполнять функцию транспорта кислорода к клеткам гемоглобин вне эритроцитов не способен.
Функциональные и нефункциональные белки плазмы.
Существуют белки плазмы, которые должны находиться в плазме и выполнять в ней функции –
их называют функциональными белками плазмы,
их дефицит в плазме может привести к патологии
(например, дефицит некоторых факторов свертывания приводит к гемофилии).
Но в плазме бывают белки, которые не выполняют в плазме никаких функций, потому их называют нефункциональными;
нефункциональные белки попадают в плазму из клеток при разрушении клеток
(то есть нефункциональные белки плазмы в норме являются внутриклеточными).
Попадание в плазму некоторых нефункциональных белков опасно:
например, попадание в кровь трипсина при панкреатите
(п.62) вследствие разрушения ПЖЖ трипсином) приводит к развитию коллапса.
Некоторые нефункциональные белки используются для диагностики
повреждений тех органов, из которых они поступили в кровь
(т.к. присутствие внутриклеточного белка данного органа в крови – результат и признак повреждения органа):
например, повышенная активность креатин/киназы в крови
может указывать на инфаркт (см. и другие примеры энзимодиагностики).
Фракции белков плазмы крови.
Белки плазмы крови делятся на две фракции:
альбумины и глобулины.
Глобулины делятся на 4 подфракции: ;1, ;2, ;, ;.
Альбуминов больше, чем глобулинов (57% против 43%),
а среди глобулинов преобладают ;-глобулины.
К фракции ;-глобулинов относятся антитела;
антитела синтезируются зрелыми В-лимфоцитами
(которые называются плазмоцитами).
Белково-пептидные гормоны, находящиеся в плазме,
синтезируются и секретируются эндокринными клетками
(например, ТТГ – гипофизом).
Остальные белки плазмы синтезируются в печени
(поэтому при повреждении печени количество БПК может снижаться).
Отклонения содержания белков плазмы от нормы.
Нормальное количество БПК – 63-83 г/л (всего – около 200г).
Снижение [БПК] называется гипо/протеин/емией,
а повышение – гипер/протеин/емией.
И снижение, и увеличение концентрации белков плазмы крови
указывает на наличие патологии в организме.
Изменение концентрации белков плазмы крови
может быть обусловлено
1) как изменением количества БПК,
2) так и изменением содержания воды в сосудах.
Например, при обезвоживании организма количество воды в сосудах меньше нормы,
и в этом случае нормальное количество БПК может сопровождаться повышенной концентрацией БПК.
Далее речь идёт об изменениях концентраций белков в плазме при нормальном количестве воды в плазме.
При воспалениях и патологии печени [альбуминов] снижается
(повышение концентрации альбуминов называется гипо/альбумин/емией),
а [глобулинов] повышается
(повышение концентрации глобулинов называется гипер/глобулин/емией).
Сочетание снижения альбуминов и повышения глобулинов называется дис/протеин/емией.
При воспалениях увеличивается концентрация ; глобулинов (;1 и ;2),
а при патологии печени увеличивается концентрация ; и ; глобулинов.
При остром воспалении нет повышения концентрации ;-глобулинов,
а при хроническом воспалении концентрация ;-глобулинов увеличена.
Снижение концентрации альбуминов наблюдается
не только при воспалениях и патологии печени, но и в других ситуациях.
Причины гипо/альбумин/емий можно разделить на три группы:
1) обусловленные снижением синтеза альбуминов (печенью),
2) обусловленные выходом альбуминов из сосудов,
3) обусловленные повышенным разрушением альбуминов.
Синтез альбуминов снижается:
1.1) при патологии печени (печень является местом синтеза большинства БПК),
1.2) при воспалительных и лихорадочных состояниях,
1.3) при дефиците «сырья» для их синтеза – аминокислот.
Дефицит аминокислот бывает:
1) при голодании (общем или белковом) или
2) при нарушении усвоения аминокислот при патологии ЖКТ
(при дефиците ферментов при патологии ПЖЖ или кишечника
или при нарушении всасывания при патологии кишечника).
Потеря белков из сосудов бывает при:
1) при повышенной проницаемости сосудов,
2) при патологии ЖКТ и почек
В последнем случае белки оказываются в моче,
присутствие белков в моче называется протеинурией;
протеинурия – признак повреждения почек,
здоровые почки не пропускают белки плазмы в мочу,
за исключение панкреатической амилазы, которой в норме в крови нет – п.62.
Повышенное разрушение альбуминов бывает при активации тканевых пептидаз.
При голодании количество белков снижается потому, что
не из чего синтезировать белки
и потому что белки используются в качестве питания
(используются для глюконеогенеза – п.33).
Ф У Н К Ц И И белков плазмы крови.
Есть функции, которые выполняются всеми БПК,
а есть функции, которые выполняются только определенными белками.
1. БПК создают онкотическое давление –
это означает, что БПК участвуют в задерживании воды в сосудах.
Другие вещества плазмы тоже участвуют в удерживании воды в сосудах,
то есть создают осмотическое давление.
Определение: онкотическое давление – это доля осмотического давления,
обусловленная белками плазмы крови.
Распределение воды между тканями и сосудами
зависит от содержания осмотически активных веществ
(то есть веществ, способных «притягивать» воду туда, где вещества находятся)
в тканях и в сосудах
(к числу осмотически активных веществ относятся белки, глюкоза, ионы):
если количество осмотически активных веществ в сосудах снижается,
то часть воды перемещается из сосудов в ткани – это приводит к отекам.
И, наоборот, при увеличении содержания осмотически активных веществ в сосудах
часть воды переходит их тканей в сосуды,
чтобы «растворять» там повышенное количество осмотически активных веществ.
При снижении количества БПК онкотическое давление снижается,
это приводит к тому, что увеличивается количество воды в тканях и возникают отеки.
2. Питательная функция белков плазмы крови.
БПК могут расщепляться до АК, которые поступают в клетки.
В клетках АК могут
1) использоваться для синтеза белков,
2) могут катаболизироваться и давать АТФ и тепло (при дефиците глюкозы и жирных кислот),
3) в печени могут превращаться в глюкозу (при гипогликемии, в печени).
Питание белками плазмы снижает их количество в крови,
приводит к гипо/протеин/емии,
проявляется отеками, функции белков плазмы крови снижаются.
Участие в регуляции рН
(в поддержании кислотно-щелочного равновесия, в [Н+]= [протонов]).
Регуляция рН имеет очень большое значение:
значительные отклонения от нормального рН крови (от слегка щелочного рН)
приводят к смерти (например, при плохо контролируемом сахарном диабете).
Белки плазмы крови способны
присоединять к себе Н+ (то есть быть акцепторами протонов – основаниями)
при избытке протонов (то есть при ацидозе) –
за счет наличия в белках радикалов, способных протонироваться:
это радикалы лизина, аргинина, гистидина (радикалы оснОвных АК).
Белки плазмы крови способны
быть источниками Н+ (то есть быть кислотами)
при дефиците Н+ (то есть при алкалозе) –
за счет наличия радикалов, способных быт источниками протонов (кислотами):
радикалы Глу и Асп с СООН группами.
Наряду с БПК существуют гемоглобиновый, фосфатный и бикарбонатный буферы (см. курс физиологии).
4. С БПК связана вязкость крови.
Чем больше [БПК], тем больше вязкость.
С п е ц и а л и з и р о в а н н ы е функции БПК.
5. Определенные белки плазмы крови участвуют в сворачивании крови –
эти БПК называют факторами свертывания крови.
Их дефицит приводит к снижению способности крови сворачиваться,
что угрожает смертью от потери крови.
При гемофилии
причиной дефицита факторов свертывания являются мутации генов
(гемофилия относится к первичным, врожденным протеинопатиям – п.57).
Причиной нарушения свертывания может быть
авитаминоз витамина К (приобретенная протеинопатия, вторичная).
6. Ряд БПК участвует в снижении свертываемости крови
(факторы противосвертывающей системы крови).
Дефицит факторов противосвёртывающей системы приводит к тромбозам.
Существуют белки плазмы, которые способны ингибировать протеазы
и тем самым спасают организм от разрушения его белков протеазами.
Ингибиторы протеазы называются анти/протеазами.
Примеры антипротеаз:
;1-антитрипсин = ;1-антипептидаза,
;2-макроглобулин.
Антитрипсин способен наряду с другими пептидазами ингибировать трипсин;
это важно при появлении трипсина в крови при панкреатите,
т.к. появление трипсина в крови приводит к коллапсу (см. п. 62).
Известно, что дефицит антипротеаз приводит к смерти
от цирроза и фиброза легких;
причиной дефицита антипротеаз являются мутации их генов.
Спасти человека в этом случае может пересадка ему печени человека с нормальными генами антипротеаз.
Концентрация в крови ряда белков плазмы крови
увеличивается при острых состояниях –
эти белки называют белками острой фазы.
Примеры белков острой фазы:
С-реактивный белок, фибриноген, антитрипсин, гаптоглобин (связывает гемоглобин при выходе гемоглобина из эритроцитов при гемолизе, это позволяет предотвратить потерю железа организмом).
9. Транспортная функция белков плазмы.
Основные транспортные белки плазмы – это альбумины.
Они связывают в плазме и транспортируют:
жирные кислоты (образующиеся при расщеплении жира в адипоцитах, в ЛПОНП и хиломикронах),
свободный билирубин (от клеток системы макрофагов к гепатоцитам, см. 118,
ионы,
лекарства (гидрофобные).
Транспортер железа называется трансферрином (он же транспортирует хром),
транспортер меди – церулоплазмином,
транспортер В12 (кобаламина) – транскобаламином,
транспортер Hb – гаптоглобином.
Есть специальные транспортеры для гидрофобных гормонов (например, для стероидов) и витаминов (А, Д).
Дефицит транспортных белков нарушает транспорт соответствующих веществ
и может привести к патологии.
Например, дефицит транспортеров витаминов
может привести к симптомам гиповитаминозов,
дефицит альбуминов:
— приводит к увеличению поступления свободного билирубина в клетки
(это нарушает состояние клеток и самочувствие) и
— требует снижения доз лекарств.
10. Участие в регуляции артериального давления:
Белки систем брадикинина и антиотензина
(пре/кинины, пре/калликреин, ангиотензиноген)
участвуют в регуляции артериального давления
(брадикинин снижает, ангиотензин увеличивает)
за счет влияния на тонус сосудов и объем плазмы крови.
Кроме этого, брадикинин увеличивает проницаемость сосудов
и вследствие этого способствует воспалительным, иммунным и аллергическим реакциям.
Ангиотензин влияет на водно-минеральный обмен:
способствует экскреции натрия, хлорида, воды и снижает экскрецию калия.
При этом сам ангиотензин является пептидом, а не белком.
Антителами и белками системы комплемента обусловлен гуморальный иммунитет.
Антитела отвечают за распознавание антигенов,
связывают их и способствуют уничтожению антигенов иммунными клетками.
Белки системы комплемента участвуют в уничтожении опасных (например, бактериальных) клеток.
Антитела образуют фракцию ;-глобулинов, секретируются плазмоцитами. Дефицит антител снижает иммунитет.
12. Апобелки липопротеинов (п.49) иногда считаются белками плазмы крови
и относятся к фракции глобулинов.
Апобелки образуются в кишечнике, как и сами хиломикроны;
в плазме в хиломикроны могут поступать апобелки из других липопротеинов, синтезированные в печени.
Функция липопротеинов в норме – транспорт липидов:
жира, холестерина, фосфолипидов и витамина Е в ткани
и (для ЛПВП) очистка тканей от избытка холестерина и тем самым предотвращение атеросклероза.
Кроме белков, растворенных в плазме, есть белки, связанные с мембранами клеток эндотелия сосудов.
Они тоже выполняют в плазме важные функции. Например, липопротеин/липаза (см. п. 49-51).
источник
Без участия печени в метаболизме белка организм может обходиться не более нескольких дней, затем наступает летальный исход. К наиболее важным функциям печени в обмене белка относят следующие.
1. Дезаминирование аминокислот.
2. Образование мочевины и извлечение аммиака из жидких сред организма.
3. Образование белков плазмы крови.
4. Взаимное превращение различных аминокислот и синтез из аминокислот других соединений.
Предварительное дезаминирование аминокислот необходимо для их использования при получении энергии и преобразования в углеводы и жиры. В небольших количествах дезаминирование осуществляется и в других тканях организма, особенно в почках, но по значимости эти процессы несопоставимы с дезаминированием аминокислот в печени.
Образование мочевины в печени помогает извлечению аммиака из жидких сред организма. Большое количество аммиака образуется в процессе дезаминирования аминокислот, дополнительное его количество постоянно образуется бактериями в кишечнике и абсорбируется в кровь. В связи с этим если в печени мочевина не образуется, то концентрация аммиака в плазме крови начинает быстро нарастать, что приводит к печеночной коме и смерти. Даже в случае резкого снижения кровотока через печень, что иногда происходит вследствие формирования шунта между воротной и полой венами, содержание аммиака в крови резко повышается с созданием условий для токсикоза.
Все основные белки плазмы крови, за исключением некоторых гамма-глобулинов, образуются клетками печени. Их количество составляет приблизительно 90% всех белков плазмы. Остальные гамма-глобулины представляют собой антитела, образуемые главным образом плазматическими клетками лимфоидной ткани. Максимальная скорость образования белков печенью составляет 15-50 г/сут, поэтому если организм теряет около половины белков плазмы, их количество может быть восстановлено в течение 1-2 нед.
Следует учитывать, что истощение белков плазмы крови является причиной быстрого наступления митотических делений гепатоцитов и увеличения размеров печени. Этот эффект сочетается с выбросом белков плазмы крови печенью, который продолжается до тех пор, пока концентрация белков в крови не вернется к нормальным значениям. При хронических заболеваниях печени (в том числе и циррозе) уровень белков в крови, особенно альбуминов, может падать до очень низких значений, что является причиной появления генерализованных отеков и асцита.
К числу наиболее важных функций печени относится ее способность синтезировать некоторые аминокислоты наряду с химическими соединениями, в состав которых включены аминокислоты. Например, в печени синтезируются так называемые заменимые аминокислоты. В процессе такого синтеза принимают участие кетокислоты, имеющие сходную химическую структуру с аминокислотами (исключая кислород в кето-положении). Аминорадикалы проходят несколько стадий трансаминирования, перемещаясь от имеющихся в надичии аминокислот в кетокислоты на место кислорода в кето-положении.
источник
Белки являются важной составной частью крови и выполняют следующие функции:
1) определяют онкотическое давление;
2) обеспечивают вязкость крови;
3) обеспечивают свертываемость крови;
4) участвуют в регуляции кислотно-основного равновесия;
5) выполняют транспортную функцию (переносят липиды, НЭЖК, металлы, билирубин, гемоглобин, гормоны, лекарственные вещества);
6) обеспечивают иммунитет (антитела, интерферон и др.);
7) питательная функция (белки являются резервом аминокислот).
Белки плазмы обычно делят на альбумины, глобулины и фибриноген.
Это простые, высокогидрофильные белки. Образуются в гепатоцитах печени. Выполняют следующие функции:
играют важную роль в поддержании коллоидно-осмотического давления крови);
транспортируют многие вещества, в том числе билирубин, катионы металлов и красок, НЭЖК, холестерин и др.;
служат богатым и быстро реализуемым резервом аминокислот.
разделены электрофоретически на подгруппы. и -глобулины вырабатываются в ретикулоэндотелиальной системе, в том числе купферовскими клетками печени.
-глобулины состоят из глико- и липопротеидов. - глобулины участвуют в транспорте различных веществ. Они имеют самую высокую электрофоре-тическую подвижность.
-глобулины состоят из глико-, липо- и металлопротеидов. Они выполняют транспортную и другие функции.
-глобулины с самой низкой электрофоретической подвижностью. К этой группе относятся большинство защитных веществ крови, многие из которых обладают ферментативной активностью. -глобулины синтезируются плазматическими клетками.
1. Собственные ферменты плазмы крови, которые участвуют в свертывании крови, растворении внутрисосудистых сгустков и т.д. Эти ферменты синтезируются в печени.
2. Клеточные ферменты освобождаются из клеток крови и клеток других тканей в результате естественного распада (лизиса). при гепатите — активность аланинаминотрансферазы, аргиназы, аспартат-сердце
Трансферрин является -глобулином. Может взаимодействовать с Сu 2+ и Zn 2+ , но главным образом связывает и переносит Fe 3+ в различные ткани.
Гаптоглобин является 2-глобулином, выполняет следующие функции:
связывает гемоглобин в соотношении 1:1, в результате образуются высокомолекулярные комплексы, которые не могут выводиться почками транспортирует витамин В12;
является естественным ингибитором катепсина В.
Церулоплазмин является 2-глобулином, выполняет следующие функции:
— является переносчиком и регулятором концентрации ионов меди в организме,
Это группа белков плазмы, содержание которых увеличивается в ответ на повреждение ткани,воспаление, опухолевый процесс. Эти белки синтезируются в печени и являются гликопротеинами. К белкам острой фазы относятся:
гаптоглобин (увеличивается в 2-3 раза, особенно при раке, ожогах, хирургических вмешательствах, воспалении);
церулоплазмин (имеет значение как антиоксидант);
трансферрин (содержание снижается);
С-реактивный белок. Отсутствует в сыворотке здорового человека, но обнаруживается при патологических состояниях, сопровождающихся некрозом
интерферон — специфический белок, появляющийся в клетках в результате проникновения в них вирусов. Он угнетает размножение вирусов в клетках.
фибриноген, основная функция которого участие в свертывании крови.
Гиперпротеинемия– увеличение общего содержания белков плазмы.рвата диарея, потеря воды организмом, а следовательно, и плазмой приводит к повышению концентрации белка в крови (относительная гиперпротеинемия). При ряде патологических состояний может наблюдатьсяабсолютная гиперпротеинемия, обусловленная увеличением уровня γ-глобулинов: например, гиперпротеинемия в результате инфекционного или токсического раздражения системы макрофагов;
Гипопротеинемия, или уменьшение общего кол-ва белка в плазме крови, наблюдается гл образом при снижении уровня альбуминов.. Содержание общего белка снижается до 30–40 г/л. Гипопротеинемия наблюдается при поражении печеночных кл (острая атрофия печени, токсический гепатит и др.). Кроме того, гипопротеинемия может возникнуть при резко увеличенной проницаемости стенок капилляров, при белковой недостаточности (поражение пищеварительного тракта, карцинома и др.).
Остаточный азот крови. Гиперазотемия, ее причины. Уремия.
остаточный азот крови (сумма всех азотсодержащих веществ крови после удаления из неё белков = Небелковый азот крови). Нормальное содержание 14,3 – 28,6 ммоль/
1) мочевина (примерно 50% 2) АКты (около 25%), 3) креатин и креатинин(7,5%; 4) полипептиды, нуклеотиды и азотистые основания (5%;
5)мочевая кислота (4%; 6) аммиак и индикан (0,5%; Индикан представляет собой калиевую или натриевую соль индоксилсерной кислоты, образующейся в печени при обезвреживании индола
Ретенционная азотемия развивается в результате недостаточного выделения с мочой азотсодержащих продуктов Она в свою очередь может быть почечной и внепочечной. При почечной ретенционной азотемии концентрация остаточного азота в крови увеличивается вследствие ослабления очистительной функции почек. 90%мочевины вместо 50%Внепочечные в свою очередь подразделяются на надпочечные и подпочечные
Продукционная азотемияразвивается при избыточном поступлении азотсодержащих продуктов в кровь, как следствие усиленного распада тканевых белков при обширных воспалениях, ранениях, ожогах, кахексии и др. Нередко наблюдаются азотемии смешанного типа.
Уреми́я— острое или хроническое самоотравление организма, обусловленное почечной недостаточностью; накопление в крови главным образом токсических продуктов азотистого обмена (азотемия), нарушения кислотно-щелочного и осмотического равновесия.
Проявления: вялость, головная боль, рвота, диарея, кожный зуд, судороги, кома и др.
Основные биохимические функции и особенности печени.
Гепатоцит имеет хорошо развитую систему эндоплазматического ретикулума ЭР как гладкую, так и шероховатую. Функции ЭР — синтез белков, (альбумины), или ферментов работающих в печени. синтезируются фосфолипиды, триглицериды и холестерол
Функции печени: 1. Пищеварительная–Она образует желчь, включающую воду (82%), желчные кислоты (12%), фосфатидилхолин (4%), холестерол (0,7%), прямой билирубин, белки.Желчь обеспечивает эмульгирование и переваривание жиров пищи, стимулирует перистальтику кишечника.
2. Экскреторнаяфункция, близка к пищеварительной – с помощью желчи выводятся билирубин, немного креатинина и мочевины, , холестерол.(в составе желчи)
3. Секреторная– печень синтез альбумина, белков свертывающей системы, липопротеинов, глюкозы, кетоновых тел, креатина.
4. Депонирующая депо гликогена, мин. в-в, особенно железо, витамины A, D, K, B12 и фолиевая кислота.
5.Метаболическая функция – поддержание метаболического гомеостаза
*Углеводный обмен.. Благодаря синтезу и распаду гликогена печень поддерживает конц-ию глюкозы в крови. Гликогена в печени30-100гр.при длительном голодании источником глюкозы яв-ся глюконеогенез из АК и глицерин. превращение гексоз (фруктозы, галактозы) в глюкозу. р-ии ПФ пути обеспечивают синтез НАДФН, необх-го для синтеза ж к-т и холестерола из глюкозы.
*Липидный обмен. Если поступает избыток глюкозы, который не используется для синтеза гликогена и других синтезов, то она превращается в липиды – холестерол и триацилглицеролы. их удаление происходит при помощи ЛПОНП.. При сильном голодании синтезируются кетоновые тела которые яв-ся альтер. ист. Е
*Белковый обмен. За 7 суток обновляются белки печени– альбумины, многие глобулины, ферменты крови, фибриноген и факторы свертывания крови. АК подвергаются реакциям с трансаминированием и дезаминированием, декарбоксилированию с образованием биогенных аминов. идет утилизация избыточного N и включение его в состав мочевины.
*Пигментный обмен. Участие превращении гидрофобного билирубина в гидрофильную форму и секреция его в желчь
6. Обезвреживающая функция — биотрансформации подвергаются: а) стероидные и тиреоидные гормоны, инсулин, адреналин, б) продукты распада гемопротеинов (билирубин), в) продукты жизнед-ти микрофлоры, всасывающихся из толстого кишечника – кадаверин (производное лизина), путресцин (производное аргинина), крезол и фенол (производное фенилаланина и тирозина) и других токсинов, г)ксенобиотики (токсины, лекарственные вещества и их метаболиты).
Взаимосвязь обмена жиров, углеводов и белков.
путем аминирования или переаминирования пировиноградная кислота, являющаяся продуктом распада углеводов, может превратиться в аминокислоту -аланин. Кроме того, пировиноградная кислота в результате дальнейших превращений дает щавелевоуксусную (СООН—СН2—СО—СООН) и a-кетоглютаровую (СООН—СН2—СН2—СО—СООН) кислоты, из которых путем реакции аминирования и переаминирования соответственно образуются аспарагиновая и глютаминовая аминокислоты.
углеводы в животном организме могут синтезироваться из продуктов окисления белков. Углеводы образуются из тех аминокислот, которые при своем дезаминировании превращаются в кетокислоты.
СВЯЗЬ МЕЖДУ ОБМЕНОМ УГЛЕВОДОВ И ЖИРОВ
Единство в обмене углеводов и жиров доказывается возникновением общих промежуточных продуктов распада. При распаде углеводов образуется пировиноградная кислота, а из нее -активная уксусная кислота -ацетил-КоА, который может быть использован в синтезе жирных кислот. Последние при своем распаде дают ацетил-КоА. Для синтеза нейтрального жира необходим кроме жирных кислот и глицерин. Глицерин также может синтезироваться из продуктов распада углеводов, а именно, из фосфоглицеринового альдегида и фосфодиоксиацетона. И наоборот, при распаде глицерина могут образовываться фосфотриозы.
СВЯЗЬ МЕЖДУ ОБМЕНОМ БЕЛКОВ И ЖИРОВ
Многие заменимые аминокислоты могут синтезироваться из промежуточных продуктов расщепления жиров. Возникающий при распаде жирных кислот цетил-КоА вступает в конденсацию с щавелевоуксусной кислотой и через цикл трикарбоновых кислот приводит к образованию a-кетоглютаровой кислоты. Кетоглютаровая кислота в результате аминирования или переаминирования переходит в глютаминовую. Глицерин, входящий в состав нейтральногo жира, окисляется в глицериновую кислоту и в дальнейшем превращается в пировиноградную, а последняя используется для синтеза заменимых аминокислот.
Использование белков для синтеза жира осуществляется через образование ацетил-КоА.
Далее ацетил-КоА может быть использован для синтеза жирных кислот. Глицерин образуется лишь за счет тех аминокислот, которые способны превращаться в пировиноградную кислоту. 
Биохимия регуляций. Основные принципы и значение. Иерархия регуляторных систем. Классификация межклеточных регуляторов. Центральная регуляция эндокринной системы: роль либеринов, статинов и тропинов.
Интеграция – это объединение элементов системы в единое целое.
Координация (соподчинение) – это подчинение менее важных элементов системы более важным элементам. Интеграция и координация – это две стороны процесса регуляции.
Внутриклеточную регуляцию (ауторегуляцию).
Дистантную регуляцию (межкеточную
Механизмы клеточной ауторегуляции
1. Компартментализация (мембранный механизм).
Роль мембран состоит в следующем:
а) мембраны делят клетки на отсеки и в каждом из них осуществляются свои процессы;
б) мембраны обеспечивают активный транспорт и регулируют потоки молекул в клетке и из клетки;
в) в мембраны встроены ферменты;
г) мембраны защищают клетку от внешних воздействий.
Воздействием на функции мембран клетка может регулировать тот или иной процесс.
2. Изменение активности ферментов.
3. Изменение количества ферм
Классификация межклеточных регуляторов
а) Гормоны – межклеточные регуляторы, доставляемые к клеткам-мишеням током крови. Вырабатываются в эндокринных железах
б) Нейрогормоны вырабатываются нервными клетками и выделяются в синаптическую щель. Нейрогормоны делятся на медиаторы и модуляторы. Медиаторы обладают непосредственным пусковым эффектом. Модуляторы изменяют эффект медиаторов. Примерами медиаторов являются ацетилхолин и норадреналин; модуляторов – -ааминомасляная кислота, дофамин.
в) Локальные гормоны – это межклеточные регуляторы, действующие на близлежащие к месту их синтеза клетки. Пример: гормоны, производные жирных кислот.
Классификация по широте действия:
а) Гормоны универсального действия действуют на все ткани организма (например, катехоламины, глюкокортикостероиды).
б) Гормоны направленного действия действуют на определенные органы-мишени (например, АКТГ действует на кору надпочечников).
Классификация по химическому строению:
а) Белково-пептидные гормоны
Полипептиды (АКТГ, глюкагон).
б) Производные аминокислот:
Катехоламины и йодтиронины — образуются из тирозина;
Ацетилхолин — образуется из серина.
Серотонин, триптамин, мелатонин — образуются из триптофана.
стероидные гормоны (гормоны коры надпочечников и половые гормоны);
производные полиненасыщенных жирных кислот (простагландины, тромбоксаны, лейкотриены).
На стимулирующие или тормозящие стимулы из ЦНС секретируются стимулирующие или ингибирующие рилизинг-факторы, которые носят название либерины или статины соответственно. Эти нейрогормоны с кровотоком достигают аденогипофиза, где стимулируют (либерины) или ингибируют (статины) биосинтез и секрецию тропных гормонов.
Тропные гормоны воздействуют на периферические железы, стимулируя выделение соответствующих периферических гормонов
Понятие о рецепторах. Механизм действия гормонов через внутриклеточные рецепторы и рецепторы плазматических мембран и вторые посредники (общая характеристика).
Рецепторы – это белковые молекулы, специфически связывающие данный гормон, в результате чего возникает какой-либо эффект.
Гормон начинает свое действие с соединения с рецептором, образуя гормон-рецепторный комплекс.
Рецепторы могут находиться внутри клетки, а также на клеточной мембране.
Механизм действия гормонов через внутриклеточные рецепторы.
Гормон проникает в клетку, связывается с рецептором. Образованный таким образом гормон-рецепторный комплекс перемещается в ядро и действует на генетический аппарат клетки. В результате меняется процесс транскрипции, а в дальнейшем, синтез белков. Таким образом, данные гормоны влияют на количество ферментов в клетке.
Механизм действия гормонов через рецепторы плазматических мембран
В этом случае гормон не проникает в клетку, а взаимодействует с рецептором на поверхности мембраны.
Первый вариант – с рецептором связан фермент, который из специфического субстрата образует второй посредник. Второй посредник далее связывается со своим рецептором в клетке. Чаще всего рецептором посредника является протеинкиназа, которая за счет фосфата АТФ, фосфорилирует белки. В результате изменяются их свойства, возникает биохимический и физиологический эффект.
Второй вариант – рецептор связан не с ферментом мембраны, а с ионным каналом. При связывании гормона с рецептором, канал открывается, ион поступает в клетку и выполняет функции второго посредника.
Хорошо изученными вторыми посредниками являются циклические нуклеотиды (цАМФ, цГМФ) и Ca 2+ .
Механизм действия гормонов через цАМФ
Когда соответствующий гормон связывается с рецептором, в мембране активируется фермент аденилатциклаза, который из АТФ образует цАМФ. цАМФ является аллостерическим активатором протеинкиназы, которая фосфорилирует белки и изменяет их свойства.
Содержание цАМФ в клетке увеличивают: глюкагон, катехоламины (через -рецепторы), антидиуретический гормон, гистамин (Н2-рецепторы), простагландин-Е, простациклин, тиреотропный гормон, АКТГ, холерный токсин.
Содержание цАМФ в клетке снижают: ацетилхолин (М-холинорецепторы), катехоламины (2-рецепторы), соматостатин, ангиотензин-II, опиаты, коклюшный токсин.
Механизм действия гормонов через Са 2+
Когда гормон связывается с рецептором, в мембране открывается кальциевый канал. В результате содержание кальция в клетке возрастает. Кальций связывается с белком клеток – кальмодулином, образуется комплекс, который может действовать непосредственно на белки, вызывая эффекты, или действовать на кальмодулин-зависимую протеинкиназу. Эта протеинкиназа фосфорилирует белки, в результате изменяются их свойства.
Са 2+ в качестве второго посредника выполняет те же функции, что и цАМФ, за исключением того, что в гладких мышцах вызывает сокращение, тромбоцитах – агрегацию.
Содержание кальция в клетке повышают: катехоламины через 1-рецепторы, ацетилхолин через М-холинорецепторы, гистамин через Н1-рецепторы, тромбоксан, ангиотензин-II.
Инсулин. Строение, образование из проинсулина, метаболизм, регуляция секреции. Влияние на обмен веществ.
Инсулин является белкого-пептидным гормоном с молекулярной массой 5700. Синтезируется в В-клетках поджелудочной железы из проинсулина. проинсулин, который транспортируется в комплекс Гольджи, далее в цистернах которого происходит так называемое созревание инсулина.
В процессе созревания из молекулы проинсулина с помощью специфических эндопептидаз вырезается C-пептид Скорость секреции инсулина зависит от концентрации глюкозы в крови: при повышении концентрации секреция инсулина увеличивается, а при снижении – уменьшается.
Молекула инсулина образована двумя полипептидными цепями, содержащими 51аминокислотный остаток: A-цепь состоит из 21 аминокислотного остатка, B-цепь образована 30 аминокислотными остатками. Полипептидные цепи соединяются двумя дисульфидными мостиками через остатки цистеина, третья дисульфидная связь расположена в A-цепи.
Секрецию инсулина также усиливают: глюкагон, секретин, холецистокинин, СТГ и пища, богатая белками.
Рецепторы для инсулина находятся на клеточной мембране, Главными мишенями для инсулина являются мышцы, печень, жировая ткань, фибробласты и лимфоциты. Головной мозг не зависит от инсулина.
усиление поглощения клетками глюкозы и других веществ;
активацию ключевых ферментов гликолиза;
увеличение интенсивности синтеза гликогена — инсулин форсирует запасание глюкозы клетками печени и мышц путём полимеризации её в гликоген;
уменьшение интенсивности глюконеогенеза — снижается образование в печени глюкозы из различных веществ
усиливает поглощение клетками аминокислот (особенно лейцина и валина);
усиливает транспорт в клетку ионов калия, а также магния и фосфата;
усиливает репликацию ДНК и биосинтез белка;
усиливает синтез жирных кислот и последующую их этерификацию — вжировой ткани и в печени инсулин способствует превращению глюкозы втриглицериды; при недостатке инсулина происходит обратное — мобилизация жиров.
Сахарный диабет. Патогенез. Нарушения обмена веществ при сахарном диабете. Определение толерантности к глюкозе при диагностике сахарного диабета.
Может быть 2 причины сахарного диабета:
Абсолютная недостаточность инсулина. В этом случае концентрация инсулина в крови ниже нормы. Это может быть связано либо с повреждением островковой ткани железы, либо с истощением запасов инсулина, либо с ускоренным его разрушением.
Относительная недостаточность возникает в результате снижения числа рецепторов к инсулину, или снижения их чувствительности.
Различают инсулинзависимый (юношеский, ювенильный) и инсулиннезависимый (стабильный) сахарный диабет.
При инсулинзависимом диабете наблюдается абсолютная недостаточность инсулина, и жизнь больных зависит от инъекции инсулина.
При инсулиннезависимом диабете наблюдается относительная недостаточность инсулина, поддержание глюкозы на нормальном уровне достигается сахаропонижающими средствами, инъекции инсулина не требуются.
источник
Печень является органом, регулирующим поступление азотистых веществ в организм и их выведение. В периферических тканях постоянно протекают реакции биосинтеза с использованием свободных аминокислот, либо выделение их в кровь при распаде тканевых белков. Несмотря на это, уровень белков и свободных аминокислот в плазме крови остаётся постоянным. Это происходит благодаря тому, что в клетках печени имеется уникальный набор ферментов, катализирующих специфические реакции обмена белков.
31.4.1. Пути использования аминокислот в печени. После приёма белковой пищи в клетки печени по воротной вене поступает большое количество аминокислот. Эти соединения могут претерпевать в печени ряд превращений, прежде чем поступить в общий кровоток. К этим реакциям относятся (рисунок 10):
а) использование аминокислот для синтеза белков;
б) трансаминирование — путь синтеза заменимых аминокислот; осуществляет также взаимосвязь обмена аминокислот с глюконеогенезом и общим путём катаболизма;
в) дезаминирование — образование α-кетокислот и аммиака;
г) синтез мочевины — путь обезвреживания аммиака (схему см. в разделе «Обмен белков»);
д) синтез небелковых азотсодержащих веществ (холина, креатина, никотинамида, нуклеотидов и т.д.).
Рисунок 10. Обмен аминокислот в печени (схема).
31.4.2. Биосинтез белков. В клетках печени синтезируются многие белки плазмы крови: альбумины (около 12 г в сутки), большинство α- и β-глобулинов, в том числе транспортные белки (ферритин, церулоплазмин, транскортин, ретинолсвязывающий белок и др.). Многие факторы свёртывания крови (фибриноген, протромбин, проконвертин, проакцелерин и др.) также синтезируются в печени.
В гепатоцитах содержатся практически все ферменты, участвующие в метаболизме липидов. Поэтому паренхиматозные клетки печени в значительной степени контролируют соотношение между потреблением и синтезом липидов в организме. Катаболизм липидов в клетках печени протекает главным образом в митохондриях и лизосомах, биосинтез — в цитозоле и эндоплазматическом ретикулуме. Ключевым метаболитом липидного обмена в печени является ацетил-КоА, главные пути образования и использования которого показаны на рисунке 5.
Рисунок 5. Образование и использование ацетил-КоА в печени.
31.3.1. Метаболизм жирных кислот в печени. Пищевые жиры в виде хиломикронов поступают в печень через систему печёночной артерии. Под действиемлипопротеинлипазы, находящейся в эндотелии капилляров, они расщепляются до жирных кислот и глицерола. Жирные кислоты, проникающие в гепатоциты, могут подвергаться окислению, модификации (укорочению или удлинению углеродной цепи, образованию двойных связей) и использоваться для синтеза эндогенных триацилглицеролов и фосфолипидов.
31.3.2. Синтез кетоновых тел. При β-окислении жирных кислот в митохондриях печени образуется ацетил-КоА, подвергающийся дальнейшему окислению в цикле Кребса. Если в клетках печени имеется дефицит оксалоацетата (например, при голодании, сахарном диабете), то происходит конденсация ацетильных групп с образованием кетоновых тел (ацетоацетат,β-гидроксибутират, ацетон). Эти вещества могут служить энергетическими субстратами в других тканях организма (скелетные мышцы, миокард, почки, при длительном голодании — головной мозг). Печень не утилизирует кетоновые тела. При избытке кетоновых тел в крови развивается метаболический ацидоз. Схема образования кетоновых тел — на рисунке 6.
Рисунок 6. Синтез кетоновых тел в митохондриях печени.
31.3.3. Образование и пути использования фосфатидной кислоты. Общим предшественником триацилглицеролов и фосфолипидов в печени является фосфатидная кислота. Она синтезируется из глицерол-3-фосфата и двух ацил-КоА — активных форм жирных кислот (рисунок 7). Глицерол-3-фосфат может образоваться либо из диоксиацетонфосфата (метаболит гликолиза), либо из свободного глицерола (продукт липолиза).
Рисунок 7. Образование фосфатидной кислоты (схема).
Для синтеза фосфолипидов (фосфатидилхолина) из фосфатидной кислоты необходимо поступление с пищей достаточного количества липотропных факторов (веществ, препятствующих развитию жировой дистрофии печени). К этим факторам относятся холин, метионин, витамин В12, фолиевая кислота и некоторые другие вещества. Фосфолипиды включаются в состав липопротеиновых комплексов и принимают участие в транспорте липидов, синтезированных в гепатоцитах, в другие ткани и органы. Недостаток липотропных факторов (при злоупотреблении жирной пищей, хроническом алкоголизме, сахарном диабете) способствует тому, что фосфатидная кислота используется для синтеза триацилглицеролов (нерастворимых в воде). Нарушение образования липопротеинов приводит к тому, что избыток ТАГ накапливается в клетках печени (жировая дистрофия) и функция этого органа нарушается. Пути использования фосфатидной кислоты в гепатоцитах и роль липотропных факторов показаны на рисунке 8.
Рисунок 8. Использование фосфатидной кислоты для синтеза триацилглицеролов и фосфолипидов. Липотропные факторы обозначены знаком * .
31.3.4. Образование холестерола. Печень является основным местом синтеза эндогенного холестерола. Это соединение необходимо для построения клеточных мембран, является предшественником желчных кислот, стероидных гормонов, витамина Д3. Первые две реакции синтеза холестерола напоминают синтез кетоновых тел, но протекают в цитоплазме гепатоцита. Ключевой фермент синтеза холестерола — β-гидрокси-β-метилглутарил-КоА-редуктаза (ГМГ-КоА-редуктаза) ингибируется избытком холестерола и желчными кислотами по принципу отрицательной обратной связи (рисунок 9).
Рисунок 9. Синтез холестерола в печени и его регуляция.
31.3.5. Образование липопротеинов. Липопротеины — белково-липидные комплексы, в состав которых входят фосфолипиды, триацилглицеролы, холестерол и его эфиры, а также белки (апопротеины). Липопротеины транспортируют нерастворимые в воде липиды к тканям. В гепатоцитах образуются два класса липопротеинов — липопротеины высокой плотности (ЛПВП) и липопротеины очень низкой плотности (ЛПОНП).
5.1. Фаза модификации. Основной тип реакций этой фазы биотрансформации — микросомальное окисление. Оно происходит при участии ферментов монооксигеназной цепи переноса электронов. Эти ферменты встроены в мембраны эндоплазматического ретикулума гепатоцитов
Источником электронов и протонов в этой цепи является НАДФН+Н + , который образуется в реакциях пентозофосфатного пути окисления глюкозы. Промежуточным акцептором Н + и е — служит флавопротеин, содержащий кофермент ФАД. Конечное звено в цепи микросомального окисления — цитохром Р-450.
Цитохром P-450 — сложный белок, хромопротеин, в качестве простетической группы содержит гем. Своё название цитохром Р-450 получил в связи с тем, что образует прочный комплекс с оксидом углерода СО, имеющий максимум поглощения при 450 нм. Цитохром Р-450 обладает низкой субстратной специфичностью. Он может взаимодействовать с большим количеством субстратов. Общее свойство всех этих субстратов — неполярность.
Цитохром Р-450 активирует молекулярный кислород и окисляемый субстрат, изменяя их электронную структуру и облегчая процесс гидроксилирования.
В этом механизме условно можно выделить 5 основных этапов:
1. Окисляемое вещество (S) образует комплекс с окисленной формой цитохрома Р-450;
2. Происходит восстановление этого комплекса электроном с НАДФН;
3. Восстановленный комплекс соединяется с молекулой O2;
4. О2 в составе комплекса присоединяет ещё один электрон с НАДФН;
5. Комплекс распадается с образованием молекулы Н2О, окисленной формы цитохрома Р-450 и гидроксилированного субстрата (S-ОН).
В отличие от митохондриальной дыхательной цепи, при переносе электронов в монооксигеназной цепи не происходит аккумулирования энергии в виде АТФ. Поэтому микросомальное окисление является свободным окислением.
В большинстве случаев гидроксилирование чужеродных веществ снижает их токсичность. Однако в ряде случаев могут образоваться продукты с цитотоксическими, мутагенными и канцерогенными свойствами.
Примеры реакций, протекающих с участием цитохрома Р-450:
источник
Тема 1. БИОЛОГИЧЕСКАЯ РОЛЬ БЕЛКОВЫХ И НЕБЕЛКОВЫХ КОМПОНЕНТОВ ПЛАЗМЫ КРОВИ.
Практическая значимость темы. Кровь является важнейшим и наиболее доступным объектом биохимического исследования. Наиболее изученные компоненты крови — это гемоглобин, альбумин, иммуноглобулины и разнообразные факторы свёртывания. При различных заболеваниях наблюдаются изменения уровня белков в плазме; эти изменения можно обнаружить при электрофорезе. Важным диагностическим признаком при некоторых патологических состояниях служит повышение активности некоторых ферментов плазмы крови. Определение содержания небелковых компонентов плазмы (глюкоза, мочевина, холестерол, билирубин и др.) также используется в диагностике заболеваний.
Цель занятия. После изучения данной темы студент должен знать состав и биологическую роль различных групп белков, небелковых азотистых компонентов (остаточного азота), безазотистых органических соединений и минеральных веществ, входящих в состав плазмы крови; уметь применять полученные знания при решении теоретических и практических задач.
Исходный уровень знаний.
- Строение и биологические функции аминокислот и белков, жирных кислот и липидов, моно- и полисахаридов.
- Участие минеральных веществ в процессах жизнедеятельности.
- Кислотно-основные свойства биологических макромолекул.
- Гидрофильные и гидрофобные свойства биологических макромолекул.
- Механизмы регуляции активности ферментов.
Общая характеристика.
Кровь — жидкая подвижная ткань, циркулирующая в замкнутой системе кровеносных сосудов, транспортирующая различные химические вещества к органам и тканям, и осуществляющая интеграцию метаболических процессов, протекающих в различных клетках.
Кровь состоит из плазмы и форменных элементов (эритроцитов, лейкоцитов и тромбоцитов). Сыворотка крови отличается от плазмы отсутствием фибриногена. 90% плазмы крови составляет вода, 10% — сухой остаток, в состав которого входят белки, небелковые азотистые компоненты (остаточный азот), безазотистые органические компоненты и минеральные вещества.
Белки плазмы крови.
Плазма крови содержит сложную многокомпонентную (более 100) смесь белков, различающихся по происхождению и функциям. Большинство белков плазмы синтезируется в печени. Иммуноглобулины и ряд других защитных белков иммунокомпетентными клетками.
Содержание общего белка в сыворотке крови здорового человека составляет 65 — 85 г/л (в плазме крови этот показатель на 2 – 4 г/л выше за счёт фибриногена).
1.2.1. Белковые фракции. При помощи высаливания белков плазмы можно выделить альбуминовую и глобулиновую фракции. В норме соотношение этих фракций составляет 1,5 – 2,5. Использование метода электрофореза на бумаге позволяет выявить 5 белковых фракций (в порядке убывания скорости миграции): альбумины, α1-, α2-, β- и γ-глобулины. При использовании более тонких методов фракционирования в каждой фракции, кроме альбуминовой, можно выделить целый ряд белков (содержание и состав белковых фракций сыворотки крови см. рисунок 1).
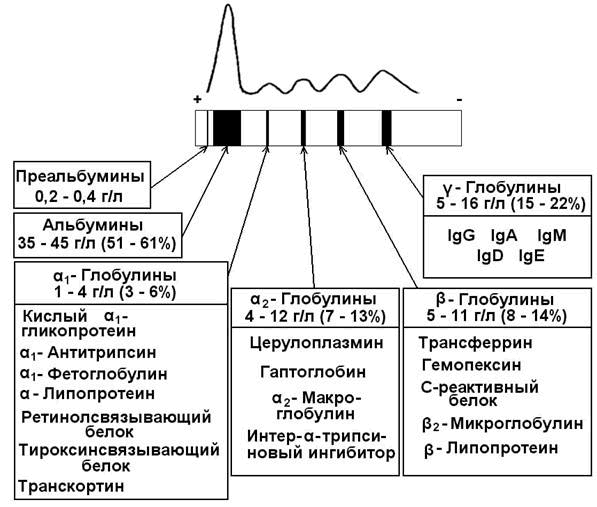
Рисунок 1. Электрофореграмма белков сыворотки крови и состав белковых фракций.
Альбумины – белки с молекулярной массой около 70000 Да. Благодаря гидрофильности и высокому содержанию в плазме играют важную роль в поддержании коллоидно-осмотического (онкотического) давления крови и регуляции обмена жидкостей между кровью и тканями. Выполняют транспортную функцию: осуществляют перенос свободных жирных кислот, желчных пигментов, стероидных гормонов, ионов Са 2+ , многих лекарств. Альбумины также служат богатым и быстро реализуемым резервом аминокислот.
α1-Глобулины:
- Кислый α1-гликопротеин (орозомукоид) – содержит до 40% углеводов, изоэлектрическая точка его находится в кислой среде (2,7). Функция этого белка до конца не установлена; известно, что на ранних стадиях воспалительного процесса орозомукоид способствует образованию коллагеновых волокон в очаге воспаления (Я.Мусил, 1985).
- α1-Антитрипсин – ингибитор ряда протеаз (трипсина, химотрипсина, калликреина, плазмина). Врождённое снижение содержания α1-антитрипсина в крови может быть фактором предрасположенности к бронхо-лёгочным заболеваниям, так как эластические волокна лёгочной ткани особенно чувствительны к действию протеолитических ферментов.
- Ретинолсвязывающий белок осуществляет транспорт жирорастворимого витамина А.
- Тироксинсвязывающий белок – связывает и транспортирует иодсодержащие гормоны щитовидной железы.
- Транскортин – связывает и транспортирует глюкокортикоидные го рмоны (кортизол, кортикостерон).
α2-Глобулины:
- Гаптоглобины (25% α2-глобулинов) – образуют стабильный комплекс с гемоглобином, появляющимся в плазме в результате внутрисосудистого гемолиза эритроцитов. Комплексы гаптоглобин-гемоглобин поглощаются клетками РЭС, где гем и белковые цепи подвергаются распаду, а железо повторно используется для синтеза гемоглобина. Тем самым предотвращается потеря железа организмом и повреждение почек гемоглобином.
- Церулоплазмин – белок, содержащий ионы меди (одна молекула церулоплазмина содержит 6-8 ионов Cu 2+ ), которые придают ему голубую окраску. Является транспортной формой ионов меди в организме. Обладает оксидазной активностью: окисляет Fe 2+ в Fe 3+ , что обеспечивает связывание железа трансферрином. Способен окислять ароматическиеамины, участвует в обмене адреналина, норадреналина, серотонина.
- Трансферрин – главный белок β-глобулиновой фракции, участвует в связывании и транспорте трёхвалентного железа в различные ткани, особенно в кроветворные. Трансферрин регулирует содержание Fe 3+ в крови, предотвращает избыточное накопление и потерю с мочой.
- Гемопексин – связывает гем и предотвращает его потерю почками. Комплекс гем-гемопексин улавливается из крови печенью.
- С-реактивный белок (С-РБ) – белок, способный преципитировать (в присутствии Са 2+ ) С-полисахарид клеточной стенки пневмококка. Биологическая роль его определяется способностью активировать фагоцитоз и ингибировать процесс агрегации тромбоцитов. У здоровых людей концентрация С-РБ в плазме ничтожно мала и стандартными методами не определяется. При остром воспалительном процессе она увеличивается более чем в 20 раз, в этом случае С-РБ обнаруживается в крови. Исследование С-РБ имеет преимущество перед другими маркерами воспалительного процесса: определением СОЭ и подсчётом числа лейкоцитов. Данный показатель более чувствителен, его увеличение происходит раньше и после выздоровления быстрее возвращается к норме.
- Иммуноглобулины (IgA, IgG, IgM, IgD, IgE) представляют собой антитела, вырабатываемые организмом в ответ на введение чужеродных веществ с антигенной активностью. Подробнее об этих белках см. 1.2.5.
1.2.2. Количественные и качественные изменения белкового состава плазмы крови. При различных патологических состояниях белковый состав плазмы крови может изменяться. Основными видами изменений являются:
- Гиперпротеинемия — увеличение содержания общего белка плазмы. Причины: потеря большого количества воды (рвота, диарея, обширные ожоги), инфекционные заболевания (за счёт увеличения количества γ-глобулинов).
- Гипопротеинемия — уменьшение содержания общего белка в плазме. Наблюдается при заболеваниях печени (вследствие нарушения синтеза белков), при заболеваниях почек (вследствие потери белков с мочой), при голодании (вследствие недостатка аминокислот для синтеза белков).
- Диспротеинемия — изменение процентного соотношения белковых фракций при нормальном содержании общего белка в плазме крови, например, снижение содержания альбуминов и увеличение содержания одной или нескольких глобулиновых фракций при различных воспалительных заболеваниях.
- Парапротеинемия — появление в плазме крови патологических иммуноглобулинов — парапротеинов, отличающихся от нормальных белков по физико-химическим свойствам и биологической активности. К таким белкам относятся, например, криоглобулины, образующие друг с другом преципитаты при температуре ниже 37° С. Парапротеины обнаруживаются в крови при макроглобулинемии Вальденстрема, при миеломной болезни (в последнем случае они могут преодолевать почечный барьер и обнаруживаться в моче как белки Бенс-Джонса). Парапротеинемия, как правило, сопровождается гиперпротеинемией.
1.2.3. Липопротеиновые фракции плазмы крови. Липопротеины — сложные соединения, осуществляющие транспорт липидов в крови. В состав их входят: гидрофобное ядро, содержащее триацилглицеролы и эфиры холестерола, и амфифильная оболочка, образованная фосфолипидами, свободным холестеролом и белками-апопротеинами (рисунок 2). В плазме крови человека содержатся следующие фракции липопротеинов:
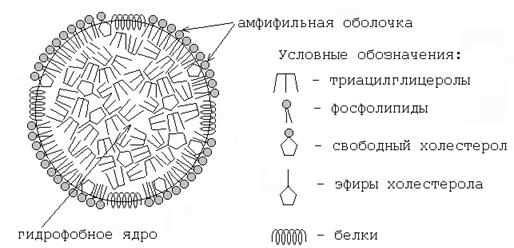
Рисунок 2. Схема строения липопротеина плазмы крови.
- Липопротеины высокой плотности или α-липопротеины, так как при электрофорезе на бумаге они движутся вместе с α-глобулинами. Содержат много белков и фосфолипидов, транспортируют холестерол из периферических тканей в печень.
- Липопротеины низкой плотности или β-липопротеины, так как при электрофорезе на бумаге они движутся вместе с β-глобулинами. Богаты холестеролом; транспортируют его из печени в периферические ткани.
- Липопротеины очень низкой плотности или пре-β-липопротеины (на электрофореграмме расположены между α- и β-глобулинами). Служат транспортной формой эндогенных триацилглицеролов, являются предшественниками липопротеинов низкой плотности.
- Хиломикроны — электрофоретически неподвижны; в крови, взятой натощак, отсутствуют. Являются транспортной формой экзогенных (пищевых) триацилглицеролов.
1.2.4. Белки острой фазы воспаления. Это белки, содержание которых увеличивается в плазме крови при остром воспалительном процессе. К ним относятся, например, следующие белки:
- гаптоглобин;
- церулоплазмин;
- С-реактивный белок;
- α1-антитрипсин;
- фибриноген (компонент свёртывающей системы крови; см. 2.2.2).
Скорость синтеза этих белков увеличивается прежде всего за счёт снижения образования альбуминов, трансферрина и альбуминов (небольшая фракция белков плазмы, обладающая наибольшей подвижностью при диск-электрофорезе, и которой соответствует полоса на электрофореграмме перед альбуминами), концентрация которых при остром воспалении снижается.
Биологическая роль белков острой фазы: а) все эти белки являются ингибиторами ферментов, освобождаемых при разрушении клеток, и предупреждают вторичное повреждение тканей; б) эти белки обладают иммунодепрессорным действием (В.Л.Доценко, 1985).
1.2.5. Защитные белки плазмы крови. К белкам, выполняющим защитную функцию, относятся иммуноглобулины и интерфероны.
Иммуноглобулины (антитела) — группа белков, вырабатываемых в ответ на попадание в организм чужеродных структур (антигенов). Они синтезируются в лимфоузлах и селезёнке лимфоцитами В. Выделяют 5 классов иммуноглобулинов — IgA, IgG, IgM, IgD, IgE.
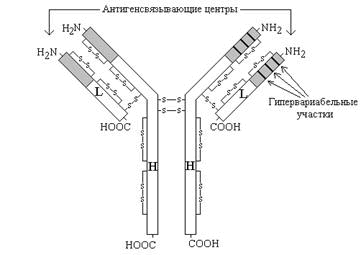
Рисунок 3.Схема строения иммуноглобулинов (серым цветом показана вариабельная область, не закрашена — константная область).
Молекулы иммуноглобулинов имеют единый план строения. Структурную единицу иммуноглобулина (мономер) образуют четыре полипептидные цепи, соединённые между собой дисульфидными связями: две тяжёлые (цепи Н) и две лёгкие (цепи L) (см. рисунок 3). IgG, IgD и IgЕ по своей структуре, как правило, являются мономерами, молекулы IgM построены из пяти мономеров, IgA состоят из двух и более структурных единиц, или являются мономерами.
Белковые цепи, входящие в состав иммуноглобулинов, можно условно разделить на специфические домены, или области, имеющие определённые структурные и функциональные особенности.
N-концевые участки как L-, так и Н-цепей называются вариабельной областью (V), так как их структура характеризуется существенными различиями у разных классов антител. Внутри вариабельного домена имеются 3 гипервариабельных участка, отличающихся наибольшим разнообразием аминокислотной последовательности. Именно вариабельная область антител ответственна за связывание антигенов по принципу комплементарности; первичная структура белковых цепей в этой области определяет специфичность антител.
С-концевые домены Н- и L-цепей обладают относительно постоянной первичной структурой в пределах каждого класса антител и называются константной областью (С). Константная область определяет свойства различных классов иммуноглобулинов, их распределение в организме, может принимать участие в запуске механизмов, вызывающих уничтожение антигенов.
Интерфероны — семейство белков, синтезируемых клетками организма в ответ на вирусную инфекцию и обладающих противовирусным эффектом. Различают несколько типов интерферонов, обладающих специфическим спектром действия: лейкоцитарный (α-интерферон), фибробластный (β-интерферон) и& иммунный (γ-интерферон). Интерфероны синтезируются и секретируются одними клетками и проявляют свой эффект, воздействуя на другие клетки, в этом отношении они подобны гормонам. Механизм действия интерферонов показан на рисунке 4.
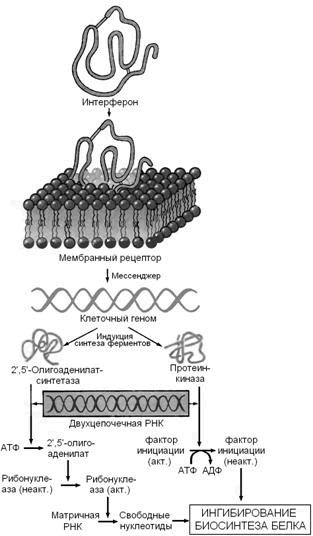
Рисунок 4.Механизм действия интерферонов (Ю.А.Овчинников, 1987).
Связываясь с клеточными рецепторами, интерфероны индуцируют синтез двух ферментов — 2′,5′-олигоаденилатсинтетазы и протеинкиназы, вероятно, за счет инициации транскрипции соответствующих генов. Оба образующихся фермента проявляют свою активность в присутствии двухцепочечных РНК, а именно такие РНК являются продуктами репликации многих вирусов или содержатся в их вирионах. Первый фермент синтезирует 2′,5′-олигоаденилаты (из АТФ), которые активируют клеточную рибонуклеазу I; второй фермент фосфорилирует фактор инициации трансляции IF2. Конечным результатом этих процессов является ингибирование биосинтеза белка и размножения вируса в инфицированной клетке (Ю.А.Овчинников, 1987).
1.2.6. Ферменты плазмы крови. Все ферменты, содержащиеся в плазме крови, можно разделить на три группы:
- секреторные ферменты — синтезируются в печени, выделяются в кровь, где выполняют свою функцию (например, факторы свёртывания крови);
- экскреторные ферменты — синтезируются в печени, в норме выделяются с желчью (например, щелочная фосфатаза), их содержание и активность в плазме крови возрастает при нарушении оттока желчи;
- индикаторные ферменты — синтезируются в различных тканях и попадают в кровь при разрушении клеток этих тканей. В разных клетках преобладают различные ферменты, поэтому при повреждении того или иного органа в крови появляются характерные для него ферменты. Это может быть использовано в диагностике заболеваний.
Например, при повреждении клеток печени (гепатит) в крови возрастает активность аланинаминотраноферазы (АЛТ), аспартатаминотрансферазы (ACT), изофермента лактатдегидрогеназы ЛДГ5, глутаматдегидрогеназы, орнитинкарбамоилтрансферазы.
При повреждении клеток миокарда (инфаркт) в крови возрастает активность аспартатаминотрансферазы (ACT), иэофермента лактатдегидрогеназы ЛДГ1, изофермента креатинкиназы MB.
При повреждении клеток поджелудочной железы (панкреатит) в крови возрастает активность трипсина, α-амилазы, липазы.
источник