| Назва | Тестовые задания для проверки остаточных знаний студентов |
| Сторінка | 4/4 |
| Дата конвертації | 09.02.2014 |
| Розмір | 0.53 Mb. |
| Тип | Документы |
mir.zavantag.com > Химия > Документы
Биохимия органов и тканей
1. К ИНДИКАТОРНЫМ ФЕРМЕНТАМ ОТНОСЯТСЯ:
- органоспецифические ферменты
- ферменты, синтезируемые в печени
- ферменты, попадающие в кровь при повреждении тканей
- ферменты, синтезируемые в любой ткани
- ферменты свертывающей системы крови
2. ФУНКЦИЯ ГАПТОГЛОБИНА:
- связывает свободный гемоглобин
- транспортирует железо
- связывает гем гемоглобина
- транспортирует медь
- транспортирует билирубин
3. КОНЕЧНЫМ ПРОДУКТОМ ПРЕОБРАЗОВАНИЯ БИЛИРУБИНА В ПЕЧЕНИ ЯВЛЯЕТСЯ:
- уробилиноген
- ди- и трипирролы
- стеркобилиноген
- моноглюкурониды билирубина
- стеркобилин
4. ОБЕЗВРЕЖИВАНИЕ БИЛИРУБИНА В ПЕЧЕНИ ПРОИСХОДИТ ПРИ УЧАСТИИ ФЕРМЕНТА:
6. ПРИ ИНФАРКТЕ МИОКАРДА В КРОВИ ПОВЫШАЕТСЯ АКТИВНОСТЬ:
- МВ — изоформ креатининфосфокиназы
- аланинаминотрансферазы
- аспартатаминотрансферазы
- щелочной фосфотазы
- верно «1» и «3»
7. СОДЕРЖАНИЕ ОБЩЕГО БИЛИРУБИНА В КРОВИ СОСТАВЛЯЕТ В НОРМЕ (МКМОЛЬ/Л):
5 .90-120
8. СНИЖЕНИЕ УСТОЙЧИВОСТИ ОРГАНИЗМА К ИНФЕКЦИОННЫМ ЗАБОЛЕВАНИЯМ МОЖЕТ БЫТЬ СВЯЗАНО С НАРУШЕНИЕМ СИНТЕЗА:
5. фибриногена
9. В НОРМЕ С МОЧОЙ ВЫВОДИТСЯ:
1. моноглюкуронид билирубина
5. стеркобилин
10. ИЗБЫТОК ЖЕЛЕЗА В РЕТИКУЛОЭНДОТЕЛИАЛЬНЫХ КЛЕТКАХ ПЕЧЕНИ И СЕЛЕЗЕНКИ ДЕПОНИРУЕТСЯ В СОСТАВЕ:
5. верно «1» и «4»
11. В ЭРИТРОЦИТАХ ГЛЮКОЗА МОЖЕТ ВКЛЮЧАТЬСЯ В СЛЕДУЮЩИЕ МЕТАБОЛИЧЕСКИЕ ПУТИ
1. аэробный распад до СО2 и Н2О
3. пентозофосфатный путь превращения глюкозы
5. верно «2» и «3»
12. К БЕЛКАМ ОСТРОЙ ФАЗЫ ОТНОСЯТСЯ
14. ПРИЧИНОЙ СЕРПОВИДНОКЛЕТОЧНОЙ АНЕМИИ ЯВЛЯЕТСЯ
1. нарушение всасывания железа
3. изменение вторичной структуры цепей глобина
4. изменение первичной структуры полипептидной цепи
5. нарушение распада гема
15. НЕПРЯМОЙ БИЛИРУБИН ПРЕВРАЩАЕТСЯ В ПРЯМОЙ БИЛИРУБИН ПУТЕМ
1. связывания с альбуминами
2. взаимодействия с ФАФС в печени
3. взаимодействия с УДФ-глюкуроновой кислотой в печени
4. связывания в кишечнике с УДФ глюкуроноволй кислотой
5. взаимодействия с ФАФС в кишечнике
16. ОСНОВНАЯ ФУНКЦИЯ ГАПТОГЛОБИНА
1. участие в реакциях острой фазы
2. участие в реакциях иммунитета
5. участие в процессах свертывания крови
17. ДИСПРОТЕИНЕМИИ ЭТО
1. увеличение концентрации общего белка в крови
2. уменьшение концентрации общего белка в крови
3. снижение в крови количества фибриногена
4. нарушение соотношения в крови белковых фракций
5. снижение в крови количества альбуинов
18. СОДЕРЖАНИЕ ОСТАТОЧНОГО АЗОТА В КРОВИ СОСТАВЛЯЕТ
5. 3,9-6,5 ммоль/л
19. ПРИ ПАРАПРОТЕИНЕМИИ В КРОВИ ПОЯВЛЯЕТСЯ
5. фетопротеин
20. ВЫБЕРИТЕ ПРАВИЛЬНОЕ УТВЕРЖДЕНИЕ
1. суточная потребность в железе равна 10-20 мг
2. после распада гема железо используется повторно
3. основная масса железа находится в цитохромах
4. железо растительных продуктов хорошо усваивается в желудочно-кишечном тракте
5. роль депо железа в организме выполняет белок трансферрин
21. ДЛЯ ГИДРОКСИЛИРОВАНИЯ ПРОЛИНА И ЛИЗИНА В КОЛЛАГЕНЕ НЕОБХОДИМ ВИТАМИН
5. рибофлавин
22. В ОБРАЗОВАНИИ ДЕСМОЗИНА УЧАСТВУЮТ
5. валин
23. КОЛЛАГЕН СИНТЕЗИРУЕТСЯ В
5. гепатоцитах
24. В ФИБРОБЛАСТАХ НА ПОЛИРИБОСОМАХ ОДНОВРЕМЕННО С СИНТЕЗОМ БЕЛКА ПРОИСХОДИТ МОДИФИКАЦИЯ АМИНОКИСЛОТНЫХ ОСТАТКОВ
1. фосфорилирование серина, треонина
2. гидроксилирование пролина, лизина
3. сульфатирование тирозина, серина
4. гликозилирование лизина, аспартата
5. метилирование
25. ГИДРОКСИЛИРОВАНИЕ ПРОЛИНА И ЛИЗИНА НЕОБХОДИМО ДЛЯ
1. формирования активного центра белка
2. образования межцепочечных связей
3. образования дисульфидных мостиков
4. поддержания отрицательного заряда белка
5. нейтрализации положительного заряда
26. МОЛЕКУЛА ПРЕПРОКОЛЛАГЕНА ПОСТУПАЕТ В
2. полость эндоплазматического ретикулума
3. межклеточное пространство
5. митохондрии
27. БЕЛКИ СОЕДИНИТЕЛЬНОЙ ТКАНИ
5. гемоглобин
28. ФИБРОНЕКТИН ПО ХИМИЧЕСКОЙ СТРУКТУРЕ ЯВЛЯЕТСЯ
5. гемопротеином
29. БИОЛОГИЧЕСКАЯ РОЛЬ ФИБРОНЕКТИНА
4. структурирует межклеточный матрикс
5. поддержание тургора
30. КОЛИЧЕСТВО ПЕПТИДНЫХ ЦЕПЕЙ В МОЛЕКУЛЕ ТРОПОКОЛЛАГЕНА
5. шесть
31. АМИНОКИСЛОТА В НАИБОЛЬШЕМ КОЛИЧЕСТВЕ ПРЕДСТАВЛЕННАЯ В КОЛЛАГЕНЕ
5. валин
32. В МОЛЕКУЛЕ КОЛЛАГЕНА ЧЕТВЕРТЬ АМИНОКИСЛОТ — ЭТО
1. пролин и гидроксипролин
5. валин и треонин
33. МАРКЕРОМ РАСПАДА КОЛЛАГЕНА ЯВЛЯЕТСЯ
5. валин
34. АМИНОКИСЛОТА, ОТСУТСТВУЮЩАЯ В СОСТАВЕ МОЛЕКУЛЫ КОЛЛАГЕНА
37. ОТНОСИТЕЛЬНАЯ ПЛОТНОСТЬ МОЧИ В НОРМЕ:
- 1,001 – 1,004 г/см 3
- 1,025 – 1,030 г/см 3
- 1,015 – 1,025 г/см3
- 1,030 – 1,033 г/см 3
- 1,050-10,65 г/см 3
38. КАЧЕСТВЕННЫМИ РЕАКЦИЯМИ НА БЕЛОК В МОЧЕ ЯВЛЯЮТСЯ:
- реакция Фелинга
- проба Геллера
- проба Гмелина
- проба с концентрированной сульфосалициловой кислотой
- верно «2» и «4»
39. КАЧЕСТВЕННЫМИ РЕАКЦИЯМИ НА ГЛЮКОЗУ В МОЧЕ ЯВЛЯЮТСЯ:
- реакция Фелинга
- проба Гмелина
- проба Ниландера
- верно «1» и «2»
- верно «1» и «3»
41. ИЗ ПЕРЕЧИСЛЕННЫХ АЗОТИСТЫХ КОМПОНЕНТОВ МОЧИ НАИБОЛЬШЕЕ КОЛИЧЕСТВО АММИАКА ВЫВОДИТСЯ В СОСТАВЕ:
- креатинина
- аммонийных солей
- индикана
- мочевины
- мочевой кислоты
42. ПРИ ПОРАЖЕНИИ ПАРЕНХИМЫ ПЕЧЕНИ В МОЧЕ ОБНАРУЖИВАЕТСЯ СЛЕДУЮЩИЙ ПАТОЛОГИЧЕСКИЙ КОМПОНЕНТ:
- кровь
- креатин
- уробилиноген
- белок
- глюкоза
43. МОЧЕВАЯ КИСЛОТА ЯВЛЯЕТСЯ КОНЕЧНЫМ ПРОДУКТОМ РАСПАДА:
- пиримидиновых оснований
- мочевины
- пуриновых оснований
- уробилиногена
- липидов
источник
В основе образования билирубина лежит разрушение железосодержащей части гемоглобина и других гемсодержащих белков и ферментов. Гем распадается до биливердина, который восстанавливается в билирубин.
Свободный билирубин токсичен, не растворяются в воде и циркулирует в крови в комплексе с альбуминами. Этот билирубин дает непрямую реакцию Ван ден Берга (после осаждения альбуминов спиртом), поэтому называется непрямым.
Непрямой билирубин, будучи связанным с альбуминами, не проходит через неповрежденные мембраны почечных клубочков и не фильтруется в мочу.
Выведение билирубина осуществляется с желчью через кишечник. Билирубин, связанный с альбуминами, доставляется кровью в печень. Билирубин легко проникает через мембраны гепатоцитов, альбумины остаются в кровотоке.
В гепатоцитах билирубин соединяется с глюкуроновой кислотой, превращаясь в билирубинмоно- и диглюкуронид («выпрямляется», прямой,связанный).
Образованные билирубинглюкурониды нетоксичны, легко растворимы. Они направляются с желчью в кишечник для выведения из организма.
Из кишечника билирубин глюкурониды частично поступают в кровоток и, находясь в крови, представляют собой фракцию прямого билирубина, который дает прямую реакцию Ван ден Берга. Прямой билирубин в отличие от непрямого легко проникает через почечные фильтры и может выделяться с мочой.
В физиологических условиях сыворотка крови содержит при мерно 25 % прямого билирубина (связанного с глюкуроновой кислотой) и 75 % непрямого билирубина (альбумин-билирубина).
Таким образом, общий билирубин крови представляет собой суммарное количество непрямого и прямого билирубина.
У здоровых людей в сыворотке крови содержится билирубина 1,7—20,5 мкмоль/литр; прямого — 0,4—5,1 мкмоль/литр.
Билирубин представляет собой конечный продукт распада гема. Основная часть (80—85%) билирубина образуется из гемоглобина и лишь небольшая часть — из других гемсодержащих белков, например цитохрома Р450. Образование билирубина происходит в клетках ретикулоэндотелиальной системы. Ежедневно образуется около 300 мг билирубина.
Преобразование гема в билирубин происходит с участием микросомального фермента гемоксигеназы, для работы которого требуются кислород и НАДФН. Расщепление порфиринового кольца происходит селективно в области метановой группы в положении а. Атом углерода, входящий в состав a-метанового мостика, окисляется до моноксида углерода, и вместо мостика образуются 2 двойные связи с молекулами кислорода, поступающими извне. Образующийся в результате этого линейный тетрапиррол по структуре является IX-aльфа-биливердином. Далее он преобразуется биливердинредуктазой, цитозольным ферментом, в IX-aльфа-билирубин. Линейный тетрапиррол такой структуры должен растворяться в воде, в то время как билирубин является жирорастворимым веществом. Растворимость в липидах определяется структурой IX-aльфа-билирубина — наличием 6 стабильных внутримолекулярных водородных связей [5]. Эти связи можно разрушить спиртом в диазореакции (Ван ден Берга), в которой неконъюгированный (непрямой) билирубин превращается в конъюгированный (прямой). In vivo стабильные водородные связи разрушаются этерификацией с помощью глюкуроновой кислоты.
Около 20% циркулирующего билирубина образуется не из гема зрелых эритроцитов, а из других источников. Небольшое количество поступает из незрелых клеток селезёнки и костного мозга. При гемолизе это количество увеличивается. Остальной билирубин образуется в печени из гемсодержащих белков, например миоглобина, цитохромов, и из других неустановленных источников. Эта фракция увеличивается при пернициозной анемии, эритропоэтической уропорфирин и при синдроме Криглера-Найяра.
Транспорт и конъюгация билирубина в печени
Неконъюгированный билирубин в плазме прочно связан с альбумином. Только очень небольшая часть билирубина способна подвергаться диализу, однако под влиянием веществ, конкурирующих с билирубином за связывание с альбумином (например, жирных кислот или органических анионов), она может увеличиваться. Это имеет важное значение у новорождённых, у которых ряд лекарств (например, сульфаниламиды и салицилаты) может облегчать диффузию билирубина в головной мозг и таким образом способствовать развитию ядерной желтухи.
Печенью выделяются многие органические анионы, в том числе жирные кислоты, жёлчные кислоты и другие компоненты жёлчи, не относящиеся к жёлчным кислотам, такие как билирубин (несмотря на его прочную связь с альбумином). Исследования показали, что билирубин отделяется от альбумина в синусоидах, диффундирует через слой воды на поверхности гепатоцита |55]. Высказанные ранее предположения о наличии рецепторов альбумина не подтвердились. Перенос билирубина через плазматическую мембрану внутрь гепатоцита осуществляется с помощью транспортных белков, например транспортного белка органических анионов [50], и/или по механизму «флип-флоп» [55]. Захват билирубина высокоэффективен благодаря его быстрому метаболизму в печени в реакции глюкуронидизации и выделению в жёлчь, а также вследствие наличия в цитозоле связывающих белков, таких как лигандины (глутатион-8-трансфераза).
Неконъюгированный билирубин представляет собой неполярное (жирорастворимое) вещество. В реакции конъюгации он превращается в полярное (водорастворимое вещество) и может благодаря этому выделяться в желчь. Эта реакция протекает с помощью микросомального фермента уридиндифосфатглюкуронилтрансферазы (УДФГТ), превращающего неконъюгированный билирубин в конъюгированный моно- и диглюкуронид билирубина. УДФГТ является одной из нескольких изоформ фермента, обеспечивающих конъюгацию эндогенных метаболитов, гормонов и нейротрансмиттеров.
Ген УДФГТ билирубина находится на 2-й паре хромосом. Структура гена сложная (рис. 12-4) [2, 54]. У всех изоформ УДФГТ постоянными компонентами являются экзоны 2—5 на 3′-конце ДНК гена. Для экспрессии гена необходимо вовлечение одного из нескольких первых экзонов. Так, для образования изоферментов билирубин-УДФГТ1*1 и 1*2 необходимо вовлечение соответственно экзонов 1А и ID. Изофермент 1*1 участвует в конъюгации практически всего билирубина, а изофермент 1*2 почти или вовсе не участвует в этом [25]. Другие экзоны (IF и 1G) кодируют изоформы фенол-УДФГТ. Таким образом, выбор одной из последовательностей экзона 1 определяет субстратную специфичность и свойства ферментов.
Дальнейшая экспрессия УДФГТ 1*1 зависит также от промоторного участка на 5′-конце, связанного с каждым из первых экзонов |6|. Промоторный участок содержит последовательность ТАТАА.
Детали строения гена важны для понимания патогенеза неконъюгированной гипербилирубинемии (синдромы Жильбера и Криглера—Найяра; см. соответствующие разделы), когда в печени содержание ферментов, ответственных за конъюгацию, снижено или они отсутствуют.
Активность УДФГТ при печёночно-клеточной желтухе поддерживается на достаточном уровне, а при холестазе даже увеличивается. У новорождённых активность УДФГТ низкая.
У человека в жёлчи билирубин представлен в основном д и глюкуронидом. Превращение билирубина в моноглюкуронид, а также в диглюкуронид происходит в одной и той же микросомальной системе глюкуронилтрансферазы [37]. При перегрузке билирубином, например при гемолизе, образуется преимущественно моноглюкуронид, а при уменьшении поступления билирубина или при индукции фермента возрастает содержание диглюкуронида.
Наиболее важное значение имеет конъюгация с глюкуроновой кислотой, однако небольшое количество билирубина конъюгируется с сульфатами, ксилозой и глюкозой; при холестазе эти процессы усиливаются [II].
В поздних стадиях холестатической или печёночно-клеточной желтухи, несмотря на высокое содержание в плазме, билирубин в моче не выявляется. Очевидно, причиной этого является образование билирубина типа III, моноконъюгированного, который ковалентно связан с альбумином [54]. Он не фильтруется в клубочках и, следовательно, не появляется в моче. Это снижает практическую значимость проб, применяемых для определения содержания билирубина в моче.
Экскреция билирубина в канальцы происходит с помощью семейства АТФ-зависимых мультиспецифичных транспортных белков для органических анионов [27]. Скорость транспорта билирубина из плазмы в жёлчь определяется этапом экскреции глюкуронида билирубина.
Жёлчные кислоты переносятся в жёлчь с помощью другого транспортного белка. Наличие разных механизмов транспорта билирубина и жёлчных кислот можно проиллюстрировать на примере синдрома Дубина—Джонсона, при котором нарушается экскреция конъюгированного билирубина, но сохраняется нормальная экскреция жёлчных кислот. Большая часть конъюгированного билирубина в жёлчи находится в смешанных мицеллах, содержащих холестерин, фосфолипиды и жёлчные кислоты. Значение аппарата Гольджи и микрофиламентов цитоскелета гепатоцитов для внутриклеточного транспорта конъюгированного билирубина пока не установлено.
Диглюкуронид билирубина, находящийся в жёлчи, водорастворим (полярная молекула), поэтому в тонкой кишке не всасывается. В толстой кишке конъюгированный билирубин подвергается гидролизу b-глюкуронидазами бактерий с образованием уробилиногенов. При бактериальном холангите часть диглюкуронида билирубина гидролизуется уже в жёлчных путях с последующей преципитацией билирубина. Этот процесс может иметь важное значение для образования билирубиновых жёлчных камней.
Распределение билирубина в тканях при желтухе
Циркулирующий билирубин, связанный с белком, с трудом проникает в тканевые жидкости с низким содержанием белка. Если количество белка в них увеличивается, желтуха становится более выраженной. Поэтому экссудаты обычно более желтушны, чем транссудаты.
Ксантохромия цереброспинальной жидкости более вероятна при менингите; классическим примером этому может служить болезнь Вейля (желтушный лептоспироз) с сочетанием желтухи и менингита.
У новорождённых может наблюдаться желтушное прокрашивание базальных ганглиев головного мозга (ядерная желтуха), обусловленное высоким уровнем неконъюгированного билирубина в крови, имеющего сродство к нервной ткани.
При желтухе содержание билирубинам цереброспинальной жидкости небольшое: одна десятая или одна сотая от уровня билирубина в сыворотке.
При выраженной желтухе внутриглазная жидкость может окрашиваться в жёлтый цвет, чем объясняется чрезвычайно редкий симптом — ксантопсия (больные видят окружающие предметы в жёлтом цвете).
При выраженной желтухе жёлчный пигмент появляется в моче, поте, семенной жидкости, молоке. Билирубин является нормальным компонентом синовиальной жидкости, может содержаться и в норме.
Цвет кожи парализованных и отёчных участков тела обычно не изменяется.
Билирубин легко связывается с эластической тканью. Она в большом количестве содержится в коже, склерах, стенке кровеносных сосудов, поэтому эти образования легко становятся желтушными. Этим же объясняется несоответствие выраженности желтухи и уровня билирубина в сыворотке в периоде выздоровления при гепатите и холестазе.
Факторы, определяющие выраженность желтухи
Даже при полной обструкции жёлчных путей выраженность желтухи может варьировать. Вслед за быстрым повышением уровень билирубина в сыворотке приблизительно через 3 нед начинает снижаться, даже если обструкция сохраняется. Выраженность желтухи зависит как от выработки жёлчного пигмента, так и от экскреторной функции почек. Скорость образования билирубина из гема может меняться; при этом возможно образование, помимо билирубина, и других продуктов, которые не вступают в диазореакцию. Билирубин, в основном неконъюгированный, может также выделяться из сыворотки слизистой оболочки кишечника.
При длительном холестазе кожа приобретает зеленоватый оттенок, вероятно вследствие отложения биливердина, не участвующего в диазореакции (Ван ден Берга), а возможно, и других пигментов.
Конъюгированный билирубин, способный растворяться в воде и проникать в жидкости тела, вызывает более выраженную желтуху, чем неконъюгированный. Внесосудистое пространство тела больше, чем внутрисосудистое. Поэтому печёночно-клеточная и холестатическая желтуха обычно более интенсивная, чем гемолитическая.
Существует 4 механизма развития желтухи.
Во-первых, возможно повышение нагрузки билирубином на гепатоциты. Во-вторых, могут нарушаться захват и перенос билирубина в гепатоцит. В-третьих, может нарушаться процесс конъюгации. И наконец, может нарушаться экскреция билирубина в жёлчь через канальцевую мембрану либо развиваться обструкция более крупных жёлчных путей.
подпечёночную, или холестатическую.
Эти типы желтухи, особенно печёночная и холестатическая, имеют во многом сходные проявления.
Надпечёночная желтуха. Уровень общего билирубина в сыворотке повышается, активность сывороточных трансаминаз и ЩФ сохраняется в пределах нормы. Билирубин представлен в основном неконъюгированной фракцией. В моче билирубин не выявляется. Этот тип желтухи развивается при гемолизе и наследственных нарушениях обмена билирубина.
Печёночная (печёночно-клеточная) желтуха (см. главы 16 и 18) обычно развивается быстро и имеет оранжевый оттенок. Больных беспокоят выраженная слабость и утомляемость. Печёночная недостаточность может быть выражена в разной степени. При лёгкой печёночной недостаточности можно выявить лишь незначительные нарушения психического статуса, более выраженная печёночная недостаточность сопровождается появлением «хлопающего» тремора, спутанности сознания и комы. Небольшая задержка жидкости может проявиться лишь увеличением массы тела, при значительной задержке жидкости появляются отёки и асцит. Вследствие нарушения синтеза печенью факторов свёртывания крови возможны кровоподтёки, как после венопункций, так и спонтанные. При биохимическом исследовании выявляют повышение активности сывороточных трансаминаз; при длительном течении заболевания возможно также снижение уровня альбумина в сыворотке.
Холестатическая желтуха (см. главу 13) развивается при нарушении поступления жёлчи в двенадцатиперстную кишку. Значительного нарушения состояния больного (помимо симптомов основного заболевания) не происходит, отмечается интенсивный зуд. Желтуха прогрессирует, в сыворотке повышаются уровень конъюгированного билирубина, активность печёночной фракции ЩФ, ГГТП, а также уровень общего холестерина и конъюгированных жёлчных кислот. Вследствие стеатореи уменьшается масса тела и нарушается всасывание витаминов А, Д, Е, К, а также кальция.
Большое значение в установлении диагноза при желтухе имеют тщательно собранный анамнез, клиническое и лабораторное обследование и биохимический и клинический анализ крови. Необходимо исследование кала, которое должно включать анализ на скрытую кровь. При исследовании мочи следует исключить повышение содержания билирубина и уробилиногена. Дополнительные методы исследования — ультразвуковое исследование (УЗИ), биопсию печени и холангиографию (эндоскопическую или чрескожную) — применяют по показаниям в зависимости от типа желтухи.
источник
141. Распад гема. Обезвреживание билирубина. Нарушения обмена билирубина—желтухи: гемолитическая, обтурационная, печеночно-клеточная. Желтуха новорожденных.
Первая реакция катаболизма гема происходит при участии NADPH-зависимого ферментативного комплекса гемоксигеназы. Ферментная сисгема локализована в мембране ЭР, в области электронтранспортных цепей микросомального окисления. Фермент катализирует расщепление связи между двумя пиррольными кольцами, содержащих винильные остатки, — таким образом, раскрывается структура кольца В ходе реакции образуются линейный тетрапир-рол — биливердин(пигмент жёлтого цвета) и монооксид углерода (СО), который получается из углерода метениловой группы. Гем индуцирует транскрипцию гена гемоксигеназы, абсолютно специфичной по отношению к тему. Ионы железа, освободившиеся при распаде гема, могут быть использованы для синтеза новых молекул гемоглобина или для синтеза других железосодержащих белков. Биливердин восстанавливается до билирубина NADPH-зависимым ферментом биливердинредуктазой. Билирубин образуется не только при распаде гемоглобина, не также при катаболизме других гемсодержащю белков, таких как цитохромы и миоглобин. При распаде 1 г гемоглобина образуется 35 мг билирубина, а в сутки у взрослого человека — примерно 250-350 мг билирубина. Дальнейший метаболизм билирубина происходит в печени.
Билирубин, образованный в клетках РЭС (селезёнки и костного мозга), плохо растворим в воде, по крови транспортируется в комплексе с белком плазмы крови альбумином. Эту форму билирубина называют неконъюгированным билирубином. Каждая молекула альбумина связывает (или даже 3) молекулы билирубина, одна из которых связана с белком более прочно (более высокое сродство), чем другие. При сдвиге рН крови в кислую сторону (повышение концентрации кетоновых тел, лактата) изменяются заряд, конформация альбумина, снижается сродство к билирубину. Поэтому билирубин, связанный с альбумином непрочно, может вытесняться из центров связывания и образовывать комплексы с коллагеном межклеточного матрикса и липидами мембран. Ряд лекарственных соединений конкурирует с билирубином за высокоаффинный, имеющий высокое сродство центр альбумина.
Поглощение билирубина паренхиматозными клетками печени. Комплекс «альбумин-билирубин», доставляемый с током крови в печеНb, на поверхности плазматической мембраны гепатоцита диссоциирует. Высвобожденный билирубин образует временный комплекс с липидами плазматической мембраны. Облегчённая диффузия билирубина в гепатоциты осуществляется двумя типами белков-переносчиков: лигандина (он транспортирует основное количество билирубина) и протеина Z. Активность поглощения билирубина гепатоцитом зависит от скорости его метаболизма в клетке. Лигандин и протеин Z обнаружены также в клетках почек и кишечника, поэтому при недостаточности функции печени они способны компенсировать ослабление процессов детоксикации в этом органе.
Конъюгация билирубина в гладком ЭР. В гладком ЭР гепатоцитов к билирубину присоединяются (реакция конъюгации) полярные группы, главным образом от глюкуроновой кислоты.Билирубин имеет 2 карбоксильные группы, поэтому может соединяться с 2 молекулами глюкуроновой кислоты, образуя хорошо растворимый в воде конъюгат — диглюкуронид билирубина (конъюгированный, или прямой, билирубин).
Донором глюкуроновой кислоты служит УДФ-глюкуронат. Специфические ферменты, УДФ-глюкуронилтрансферазы (уридиндифосфоглюкуронилтрансферазы) катализируют образование моно- и диглюкуронидов билирубина.
Секреция конъюгированного билирубина в жёлчь идёт по механизму активного транспорта, т.е. против градиента концентрации. Активный транспорт является, вероятно, ско-рость-лимитирующей стадией всего процесса метаболизма билирубина в печени. В норме диглюкуронид билирубина — главная форма экскреции билирубина в жёлчь, однако не исключается присутствие небольшого количества моноглюкуронида. Транспорт конъюгирован-ного билирубина из печени в жёлчь активируется теми же лекарствами, которые способны индуцировать конъюгацию билирубина. Таким образом, можно сказать, что скорость конъюгации билирубина и активный транспорт билирубинглюкуронида из гепатоцитов в жёлчь строго взаимосвязаны В кишечнике поступившие билирубинглюкурониды гидролизуются специфическими бактериальными ерментами β-глюкуронидазами, которые гидролизуют связь между билирубином и остатком глюкуроновой кислоты. Освободившийся в ходе этой реакции билирубин под действием кишечной микрофлоры восстанавливается с образованием группы бесцветных тет-рапиррольных соединений – уробилиногенов В подвздошной и толстой кишках небольшая часть уробилиногенов снова всасывается, попадает с кровью воротной вены в печеНb. Основная часть уробилиногена из печени в составе жёлчи выводится в кишечник и выделяется с фекалиями из организма, часть уробилиногена из печени поступает в кровь и удаляется с мочой в форме уробилина. В норме большая часть бесцветных уробилиногенов, образующихся в толстой кишке, под действием кишечной микрофлоры окисляется в прямой кишке до пигмента коричневого цвета уробилина и удаляется с фекалиями. Цвет фекалий обусловлен присутствием уробилина.
Причинами гипербилирубинемии могут быть увеличение образования билирубина, превышающее способность печени экскретировать его, или повреждение печени, приводящее к нарушению секреции билирубина в жёлчь в нормальных количествах. Гипербилирубинемию отмечают также при закупорке желчевыводящих протоков печени. Во всех случаях содержание билирубина в крови повышается. При достижении определённой концентрации он диффундирует в ткани, окрашивая их в жёлтый цвет. Пожелтение тканей из-за отложения в них билирубина называют желтухой. Клинически желтуха может не проявляться до тех пор, пока концентрация билирубина в плазме крови не превысит верхний предел нормы более чем в 2,5 раза, т.е. не станет выше 50 мкмоль/л.
Гемолитическая (надпечёночная) желтуха. Известно, что способность печени образовывать глюкурониды и выделять их в жёлчь в 3-4 раза превышает их образование в физиологических условиях. Гемолитическая (надпечёночная) желтуха — результат интенсивного гемолиза эритроцитов. Она обусловлена чрезмерным образованием билирубина, превышающим способность печени к его выведению. Гемолитическая желтуха развивается при исчерпании резервных возможностей печени. Основная причина надпечёночной желтухи — наследственные или приобретённые гемолитические анемии. При гемолитических анемиях, вызванных сепсисом, лучевой болезнью, дефицитом глюкозо-6-фосфатдегидрогеназы эритроцитов, та-лассемией, переливанием несовместимых групп крови, отравлением сульфаниламидами, количество освобождающегося из эритроцитов гемоглобина за сутки может доходить до 45 г (при норме 6,25 г), что значительно увеличивает образование билирубина. Гипербилирубинемия у больных гемолитической желтухой обусловлена значительным повышением (103 — 171 мкмоль/л) в крови концентрации альбуминсвязанного не-конъюгированного билирубина (непрямой билирубин). Образование в печени и поступление в кишечник больших количеств билирубинглюкуронидов (прямой билирубин) ведёт к усиленному образованию и выделению с калом и мочой уробилиногенов и более интенсивной их окраски. Один из главных признаков гемолитической желтухи — повышение содержания в крови не-конъюгированиого (непрямого) билирубина. Это позволяет легко отличить её от механической (подпечёночной) и печёночно-клеточной (печёночной) желтух. Неконъюгированный билирубин токсичен. Гидрофобный, липофильный неконъюгирован-ный билирубин, легко растворяясь в липидах мембраны и проникая вследствие этого в митохондрии, разобщает в них дыхание и окислительное фосфорилирование, нарушает синтез белка, поток ионов калия через мембрану клетки и органелл. Это отрицательно сказывается а состоянии ЦНС, вызывая у больных ряд характерных неврологических симптомов.
Желтуха новорождённых. Частая разновидность гемолитической желтухи новорождённых — «физиологическая желтуха», наблюдающаяся в первые дни жизни ребёнка. Причиной повышения концентрации непрямого билирубина в крови служит ускоренный гемолиз и недостаточность функции белков и ферментов печени, ответственных за поглощение, конъюгацию и секрецию прямого билирубина. У новорождённых не только снижена активность УДФ-глюкуронилтрансферазы, но и, по-видимому, недостаточно активно происходит синтез второго субстрата реакции конъюгации УДФ-глюкуроната. Известно, что УДФ-глюкуронилтрансфераза — индуцируемый фермент. Одно из неприятных осложнений «физиологической желтухи» — билирубиновая энцефалопатия. Когда концентрация неконъюгированного билирубина превышает 340 мкмоль/л, он проходит через гематоэнцефалический барьер головного мозга и вызывает его поражение.
Печёночно-клеточная (печёночная) желтуха. Печёночно-клеточная (печёночная) желтуха обусловлена повреждением гепатоцитов и жёлчных капилляров, например, при острых вирусных инфекциях, хроническом и токсических гепатитах.Причина повышения концентрации билирубина в крови — поражение и некроз части печёночных клеток. Происходит задержка билирубина в печени, чему способствует резкое ослабление метаболических процессов в поражённых гепатоцитах, которые теряют способность нормально выполнять различные биохимические и физиологические процессы, в частности переводить конъюгированный (прямой) билирубин из клеток в жёлчь против градиента концентрации. Для печёночно-клеточной желтухи характерно то, что вместо преобладающих в норме диглюкуронидов билирубина в поражённой печёночной клетке образуются главным образом моноглюкурониды В результате деструкции печёночной паренхимы образующийся прямой билирубин частично попадает в большой круг кровообращения, что ведёт к желтухе. Экскреция жёлчи также нарушена. Билирубина в кишечник попадает меньше, чем в норме. При печёночно-клеточной желтухе повышается концентрация в крови как общего билирубина, так и обеих его фракций — неконъюгированного (непрямого) и конъюгированного (прямого). Так как в кишечник поступает меньше билирубинглюкуронида, то и количество образующегося уробилиногена также снижено. Поэтому кал гипохоличный, т.е. менее окрашенный. Моча, наоборот, имеет более интенсивную окраску за счёт присутствия там не только уробилинов, но и конъюгированного билирубина, который хорошо растворим в воде и экскретируется с мочой.
Механическая, или обтурационная (подпечёночная) желтуха. Механическая, или обтурационная (подпечёночная), желтуха развивается при нарушении желчеотделения в двенадцатиперстную кишку. Это может быть вызвано закупоркой жёлчных протоков, например при желчнокаменной болезни, опухолью поджелудочной железы, жёлчного пузыря, печени, двенадцатиперстной кишки, хроническим воспалением поджелудочной железы или послеоперационным сужением общего жёлчного протока При полной закупорке общего жёлчного протока конъюгированный билирубин в составе желчи не поступает в кишечник, хотя гепатоциты продолжают его вырабатывать. Поскольку билирубин в кишечник не попадает, продуктов его катаболизма уробилиногенов в моче и кале нет. Кал обесцвечен. Так как нормальные пути экскреции билирубина заблокированы, происходит его утечка в кровь, поэтому в крови больных повышена концентрация конъюгированного билирубина. Растворимый билирубин экскретируется с мочой, придавая ей насыщенный оранжево-коричневый цвет.
источник
11.1 Расставьте цифры в порядке поступления холестерина из кишечника в печень.
Гидролиз эфира холестерина пищи.
Образование смешанных мицелл.
Захват печенью остаточных хиломикронов.
Образование остаточных хиломикронов.
11.2 Составьте схему синтеза гликогена в печени, используя перечисленные ферменты:
11.3 Перечислите последовательность событий, протекающих в гепатоцитах под влиянием глюкагона:
аденилатциклаза неактивная→аденилатциклаза активная;
протеинкиназа неактивная→протеинкиназа активная;
фосфорилаза неактивная→фосфорилаза активная;
11.4 Расставьте цифры в порядке поступления холестерина из печени в периферические ткани:
синтез холестерина и его жиров;
11.5 Непрямой билирубин образуется при распаде …
11.6 Прямой билирубин образуется в печени за счет связывания …
11.7 Перечислите свойства прямого и непрямого билирубина, а также их общие свойства.
3. Легко выводится из организма.
4. Концентрация увеличивается при гемолитической желтухе.
5. Концентрация увеличивается при обтурационной желтухе.
6. Транспортируется кровью в комплексе с альбуминами.
7. Является связанным с глюкуроновой кислотой.
11.8 Найдите положения, соответствующие аэробному окислению лактата и глюконеогенезу из лактата в печени:
А – глюконеогенез из лактата;
1. Снижение в клетке соотношения АТФ/АДФ влияет на скорость процесса.
2. Накопление цитрата увеличивает скорость.
3. Сопровождается синтезом 18 молекул АТФ.
4. Затрачивается 6 АТФ на активацию процесса.
5. Накопление НАДН2ингибирует процесс.
6. Регуляторный фермент пируваткарбоксилаза.
11.9 Детоксикация этилового спирта в печени осуществляется следующими путями:
11.10 Выберите белки, синтезируемые только в печени:
11.11 При длительном употреблении алкоголя происходят следующие отклонения:
усиление распада гликогена;
повышение энергетического метаболизма;
11.12 В немикросомальном окислении ксенобиотиков принимают участие следующие ферменты:
11.13 Энергозависимыми являются следующие реакции конъюгации:
11.14 По биохимическому принципу ксенобиотики классифицируются на:
блокаторы функциональных групп белков и коферментов.
11.15 Для микросомального обезвреживания токсических веществ характерны следующие реакции:
11.16 В процессе немикросомального окисления ксенобиотиков принимают участие следующие ферменты:
11.17 Путем микросомального окисления в печени происходит:
синтез холестерина и стероидных гормонов;
синтез ненасыщенных жирных кислот;
гидроксилирование биогенных аминов.
11.18 На рисунке изображено:
 О2
О2
 2Н + 2ē 2ē 2ēRH
2Н + 2ē 2ē 2ēRH
Н

 АДФН2ФП 1 цит. Р450
АДФН2ФП 1 цит. Р450
 ROH
ROH

 2Н + HOH
2Н + HOH
редуктазная цепь окисления ксенобиотиков;
дыхательная цепь наружной мембраны митохондрий;
монооксигеназная цепь ксенобиотиков;
дахательная цепь внутренней мембраны митохондрий.
11.19 На рисунке изображено:
 О2
О2
 2Н + 2ē 2ē 2ē 2ēRH
2Н + 2ē 2ē 2ē 2ēRH
Н


 АДН2ФП 2 цит.В5 цит. Р450
АДН2ФП 2 цит.В5 цит. Р450
 ROH
ROH

 2Н + HOH
2Н + HOH
монооксигеназная цепь окисления;
редуктазная цепь окисления ксенобиотиков;
дыхательная цепь внутренней мембраны митохондрий;
конечный путь преобразования билирубина.
11.20 При длительном введении алкоголя в организме происходят следующие отклонения:
обладает абсолютной специфичностью, так как действует только на определенные субстраты;
мало специфичен, так как действует на большинство гидрофобных субстратов;
принимает протоны и электроны от любых субстратов;
не обладает аутооксидабельностью.
11.22 В печени глюкозо-6-фосфат выполняет следующие функции:
является субстратом для пентозного пути окисления;
активирует фосфоролиз гликогена;
участвует в синтезе гликогена.
11.23 Выберите реакции синтеза липидов, протекающие только в печени:
11.24 В печени НАДФН2используется для синтеза:
11.25 В печени протекают следующие реакции метаболизма липидов:
синтез и окисление жирных кислот;
синтез и окисление кетоновых тел;
11.26 Найдите отличия обмена гликогена в печени от использования его мышцами:
в печени отсутствует глюкозо-6-фосфатаза;
гликоген печени используется только на нужды печени;
в мышцах идет цикл Кори, а в печени нет;
гликоген печени используется на нужды всего организма.
11.27 Выберите ферменты, проявляющие наибольшую активность в печени:
креатинфосфокиназа ММ и МВ;
11.28 При обтурационной желтухе:
нарушен процесс желчевыделения;
нарушен процесс транспорта непрямого билирубина;
в крови увеличен прямой и непрямой билирубин;
нарушен процесс конъюгации с глюкуроновой кислотой;
в кале отсутствует стеркобилиноген.
11.29 Нормальные показатели пигментного обмена:
в крови содержится 75% непрямого и 25% прямого билирубина;
в моче содержится билирубин;
в моче содержится стеркобилиноген;
в кале отсутствуют желчные пигменты.
11.30 При паренхиматозной желтухе:
нарушена экскреция прямого билирубина в желчные капилляры;
усилен гемолиз эритроцитов;
в крови и моче появляется уробилиноген;
в кале увеличивается количество стеркобилиногена;
нарушена активность УДФ-глюкуронил-трансферазы.
связан с глюкуроновой кислотой;
адсорбирован на белках сыворотки крови;
ковалентно связан с альбуминами сыворотки крови;
11.32 При полном удалении печени, концентрация каких из перечисленных веществ в крови уменьшается?
транспортируется альбуминами крови;
связан с глюкуроновой кислотой;
связан в печени с желчными кислотами;
дает цветную реакцию с диазореактивом Эрлиха.
11.34 Что наблюдается при активации глюкуронилтрансферазы в гепатоцитах?
происходит уменьшение количества прямого билирубина в крови;
происходит увеличение количества прямого билирубина в крови;
происходит увеличение количества непрямого билирубина в крови;
не изменяется соотношение прямого и непрямого билирубина в крови.
11.35 При гемолитической желтухе:
происходит усиленный распад гемоглобина;
в крови увеличено содержание прямого билирубина;
в моче появляется билирубин;
в крови резко увеличен непрямой билирубин;
в моче отсутствует билирубин;
11.36 Выберите конечный продукт преобразования билирубина в печени:
источник
За сутки у человека распадается около 9 г гемопротеинов, в основном это гемоглобин эритроцитов.
Эритроциты в норме живут 90-120 дней, после чего лизируются в клетках ретикулоэндотелиальной системы – макрофагах селезенки (главным образом), купферовских клетках печени и макрофагах костного мозга. При разрушении эритроцитов в кровеносном русле высвобождаемый гемоглобин образует комплекс с белком-переносчиком гаптоглобином (фракция α2-глобулинов крови) и также переносится в клетки РЭС селезенки, печени и костного мозга.
В клетках РЭС гем в составе гемоглобина окисляется молекулярным кислородом. В реакциях последовательно происходит разрыв метинового мостика между 1-м и 2-м пиррольными кольцами гема с их восстановлением, отщеплением железа и белковой части и образованием оранжевого пигмента билирубина. Высвобождаемое железо может либо запасаться в клетке в комплексе с ферритином, либо выделяться наружу и связываться с трансферрином.
Билирубин – токсичное, жирорастворимое вещество, способное разобщать окислительное фосфорилирование в клетках. Особенно чувствительны к нему клетки нервной ткани.
Из клеток ретикуло-эндотелиальной системы билирубин попадает в кровь. Здесь он находится в комплексе с альбумином плазмы, в гораздо меньшем количестве – в комплексах с металлами, аминокислотами, пептидами и другими малыми молекулами. Образование таких комплексов не позволяет выделяться билирубину с мочой. Билирубин в комплексе с альбумином называется свободный (неконъюгированный) или непрямой билирубин.
Из сосудистого русла в гепатоциты билирубин попадает с помощью белка-переносчика (транспортный белок органических анионов) или по механизму флип-флоп. Далее при участии цитозольного связывающего белка лигандина (Y-протеин) билирубин транспортируется в ЭПР, где протекает реакция связывания билирубина с УДФ-глюкуроновой кислотой, при этом образуются моно — и диглюкурониды . Кроме глюкуроновой кислоты, в реакцию конъюгации могут вступать сульфаты, фосфаты, глюкозиды.
Билирубин-глюкуронид получил название связанный (конъюгированный) или прямой билирубин.
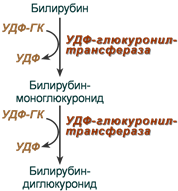
После образования билирубин-глюкурониды АТФ-зависимым переносчиком секретируются в желчные протоки и далее в кишечник, где при участии бактериальной β-глюкуронидазы превращаются в свободный билирубин. Одновременно, даже в норме (особенно у взрослых), некоторое количество билирубин-глюкуронидов может попадать из желчи в кровь по межклеточным щелям.
Таким образом, в плазме крови обычно присутствуют две формы билирубина: свободный (непрямой) , попадающий сюда из клеток РЭС (80% и более всего количества), и связанный (прямой) , попадающий из желчных протоков (в норме не более 20%).
Термины «связанный«, «конъюгированный«, «свободный«, «несвязанный» отражают взаимодействие билирубина и глюкуроновой кислоты (но не билирубина и альбумина!).
Термины «прямой» и «непрямой» введены, исходя из возможности химической реакции билирубина с диазореактивом Эрлиха. Связанный билирубин реагирует с реактивом напрямую , без добавления дополнительных реагентов, т.к. является водорастворимым. Несвязанный (жирорастворимый) билирубин требует добавочных реактивов, реагирует не прямо.
В кишечнике билирубин подвергается восстановлению под действием микрофлоры до мезобилирубина и мезобилиногена ( уробилиногена ). Часть уробилиногена всасывается и с кровью портальной вены попадает в печень, где либо распадается до моно-, ди- и трипирролов, либо окисляется до билирубина и снова экскретируется. При этом при здоровой печени в общий круг кровообращения и в мочу мезобилирубин и уробилиноген не попадают, а полностью задерживаются гепатоцитами.
Оставшаяся в кишечнике часть пигментов ферментами бактериальной флоры толстого кишечника восстанавливается до стеркобилиногена . Далее
- малая часть стеркобилиногена может всасываться и катаболизировать в печени, подобно уробилиногену,
- незначительное количество стеркобилиногена через геморроидальные вены попадает в большой круг кровообращения, отсюда в почки и в мочу. После окисления на воздухе из стеркобилиногена образуется стеркобилин мочи,
- однако основное количество стеркобилиногена достигает нижних отделов толстого кишечника и выделяется. В прямой кишке и на воздухе стеркобилиноген окисляется в стеркобилин , окрашивая кал,
- аналогично уробилиноген, появляющийся в моче при патологии печени, окисляется в уробилин .
источник
11.1 Расставьте цифры в порядке поступления холестерина из кишечника в печень.
Гидролиз эфира холестерина пищи.
Образование смешанных мицелл.
Захват печенью остаточных хиломикронов.
Образование остаточных хиломикронов.
11.2 Составьте схему синтеза гликогена в печени, используя перечисленные ферменты:
11.3 Перечислите последовательность событий, протекающих в гепатоцитах под влиянием глюкагона:
аденилатциклаза неактивная→аденилатциклаза активная;
протеинкиназа неактивная→протеинкиназа активная;
фосфорилаза неактивная→фосфорилаза активная;
11.4 Расставьте цифры в порядке поступления холестерина из печени в периферические ткани:
синтез холестерина и его жиров;
11.5 Непрямой билирубин образуется при распаде …
11.6 Прямой билирубин образуется в печени за счет связывания …
11.7 Перечислите свойства прямого и непрямого билирубина, а также их общие свойства.
3. Легко выводится из организма.
4. Концентрация увеличивается при гемолитической желтухе.
5. Концентрация увеличивается при обтурационной желтухе.
6. Транспортируется кровью в комплексе с альбуминами.
7. Является связанным с глюкуроновой кислотой.
11.8 Найдите положения, соответствующие аэробному окислению лактата и глюконеогенезу из лактата в печени:
А – глюконеогенез из лактата;
1. Снижение в клетке соотношения АТФ/АДФ влияет на скорость процесса.
2. Накопление цитрата увеличивает скорость.
3. Сопровождается синтезом 18 молекул АТФ.
4. Затрачивается 6 АТФ на активацию процесса.
5. Накопление НАДН2ингибирует процесс.
6. Регуляторный фермент пируваткарбоксилаза.
11.9 Детоксикация этилового спирта в печени осуществляется следующими путями:
11.10 Выберите белки, синтезируемые только в печени:
11.11 При длительном употреблении алкоголя происходят следующие отклонения:
усиление распада гликогена;
повышение энергетического метаболизма;
11.12 В немикросомальном окислении ксенобиотиков принимают участие следующие ферменты:
11.13 Энергозависимыми являются следующие реакции конъюгации:
11.14 По биохимическому принципу ксенобиотики классифицируются на:
блокаторы функциональных групп белков и коферментов.
11.15 Для микросомального обезвреживания токсических веществ характерны следующие реакции:
11.16 В процессе немикросомального окисления ксенобиотиков принимают участие следующие ферменты:
11.17 Путем микросомального окисления в печени происходит:
синтез холестерина и стероидных гормонов;
синтез ненасыщенных жирных кислот;
гидроксилирование биогенных аминов.
11.18 На рисунке изображено:
 О2
О2
 2Н + 2ē 2ē 2ēRH
2Н + 2ē 2ē 2ēRH


 НАДФН2ФП 1 цит. Р450
НАДФН2ФП 1 цит. Р450
 ROH
ROH

 2Н + HOH
2Н + HOH
редуктазная цепь окисления ксенобиотиков;
дыхательная цепь наружной мембраны митохондрий;
монооксигеназная цепь ксенобиотиков;
дахательная цепь внутренней мембраны митохондрий.
11.19 На рисунке изображено:
 О2
О2
 2Н + 2ē 2ē 2ē 2ēRH
2Н + 2ē 2ē 2ē 2ēRH



 НАДН2ФП 2 цит.В5 цит. Р450
НАДН2ФП 2 цит.В5 цит. Р450
 ROH
ROH

 2Н + HOH
2Н + HOH
монооксигеназная цепь окисления;
редуктазная цепь окисления ксенобиотиков;
дыхательная цепь внутренней мембраны митохондрий;
конечный путь преобразования билирубина.
11.20 При длительном введении алкоголя в организме происходят следующие отклонения:
обладает абсолютной специфичностью, так как действует только на определенные субстраты;
мало специфичен, так как действует на большинство гидрофобных субстратов;
принимает протоны и электроны от любых субстратов;
не обладает аутооксидабельностью.
11.22 В печени глюкозо-6-фосфат выполняет следующие функции:
является субстратом для пентозного пути окисления;
активирует фосфоролиз гликогена;
участвует в синтезе гликогена.
11.23 Выберите реакции синтеза липидов, протекающие только в печени:
11.24 В печени НАДФН2используется для синтеза:
11.25 В печени протекают следующие реакции метаболизма липидов:
синтез и окисление жирных кислот;
синтез и окисление кетоновых тел;
11.26 Найдите отличия обмена гликогена в печени от использования его мышцами:
в печени отсутствует глюкозо-6-фосфатаза;
гликоген печени используется только на нужды печени;
в мышцах идет цикл Кори, а в печени нет;
гликоген печени используется на нужды всего организма.
11.27 Выберите ферменты, проявляющие наибольшую активность в печени:
креатинфосфокиназа ММ и МВ;
11.28 При обтурационной желтухе:
нарушен процесс желчевыделения;
нарушен процесс транспорта непрямого билирубина;
в крови увеличен прямой и непрямой билирубин;
нарушен процесс конъюгации с глюкуроновой кислотой;
в кале отсутствует стеркобилиноген.
11.29 Нормальные показатели пигментного обмена:
в крови содержится 75% непрямого и 25% прямого билирубина;
в моче содержится билирубин;
в моче содержится стеркобилиноген;
в кале отсутствуют желчные пигменты.
11.30 При паренхиматозной желтухе:
нарушена экскреция прямого билирубина в желчные капилляры;
усилен гемолиз эритроцитов;
в крови и моче появляется уробилиноген;
в кале увеличивается количество стеркобилиногена;
нарушена активность УДФ-глюкуронил-трансферазы.
связан с глюкуроновой кислотой;
адсорбирован на белках сыворотки крови;
ковалентно связан с альбуминами сыворотки крови;
11.32 При полном удалении печени, концентрация каких из перечисленных веществ в крови уменьшается?
транспортируется альбуминами крови;
связан с глюкуроновой кислотой;
связан в печени с желчными кислотами;
дает цветную реакцию с диазореактивом Эрлиха.
11.34 Что наблюдается при активации глюкуронилтрансферазы в гепатоцитах?
происходит уменьшение количества прямого билирубина в крови;
происходит увеличение количества прямого билирубина в крови;
происходит увеличение количества непрямого билирубина в крови;
не изменяется соотношение прямого и непрямого билирубина в крови.
11.35 При гемолитической желтухе:
происходит усиленный распад гемоглобина;
в крови увеличено содержание прямого билирубина;
в моче появляется билирубин;
в крови резко увеличен непрямой билирубин;
в моче отсутствует билирубин;
11.36 Выберите конечный продукт преобразования билирубина в печени:
источник