Печень- это самый крупный наш орган, его масса составляет от 3 до 5% массы тела. Основную массу органа составляют клетки гепатоциты. Это название часто встречается, когда речь заходит о функциях и болезнях печени, поэтому запомним его. Гепатоциты специально приспособлены для синтеза, преобразования и хранения множества различных веществ, которые поступают из крови – и в большинстве случаев туда же возвращаются. Вся наша кровь протекает через печень; она наполняет многочисленные печеночные сосуды и специальные полости, а вокруг них сплошным тонким слоем расположились гепатоциты. Такая структура облегчает обмен веществ между печеночными клетками и кровью.
Печень – депо крови
В печени очень много крови, но не вся она «проточная». Довольно значительный ее объем находится в резерве. При большой потере крови сосуды печени сжимаются и выталкивают свои запасы в общее кровеносное русло, спасая человека от шока.
Печень выделяет желчь
Выделение желчи – одна из важнейших пищеварительных функций печени. Из печеночных клеток желчь поступает в желчные капилляры, которые объединяются в проток, впадающий в двенадцатиперстную кишку. Желчь вместе с пищеварительными ферментами разлагает жир на составляющие и облегчает его всасывание в кишечнике.
Печень синтезирует и разрушает жиры
Клетки печени синтезируют некоторые жирные кислоты и их производные, необходимые организму. Правда, есть среди этих соединений и те, которые многие считают вредными, – это липопротеиды низкой плотности (ЛПНП) и холестерин, избыток которых образует атеросклеротические бляшки в сосудах. Но не спешите ругать печень: мы не можем обойтись без этих веществ. Холестерин – непременный компонент мембран эритроцитов (красных кровяных телец), а доставляют его к месту образования эритроцитов именно ЛПНП. Если холестерина слишком много, эритроциты теряют эластичность и с трудом протискиваются сквозь тонкие капилляры. Люди думают, что у них проблемы с кровообращением, а у них печень не в порядке. Здоровая печень мешает образованию атеросклеротических бляшек, ее клетки извлекают из крови избыток ЛПНП, холестерина и других жиров и разрушают их.
Печень синтезирует белки плазмы крови.
Почти половина белка, который синтезирует за сутки наш организм, образуется в печени. Самые важные среди них – белки плазмы крови, прежде всего альбумин. На его долю приходится 50% всех белков, создаваемых печенью. В плазме крови должна быть определенная концентрация белков, и поддерживает ее именно альбумин. Кроме того, он связывает и переносит многие вещества: гормоны, жирные кислоты, микроэлементы. Помимо альбумина, гепатоциты синтезируют белки свертывания крови, препятствующие образованию тромбов, а также многие другие. Когда белки состарятся, их распад происходит в печени.
В печени образуется мочевина
Белки в нашем кишечнике расщепляются на аминокислоты. Часть из них находит применение в организме, а остальные нужно удалить, потому что запасать их организм не может. Распад ненужных аминокислот происходит в печени, при этом образуется токсичный аммиак. Но печень не позволяет организму отравиться и сразу преобразует аммиак в растворимую мочевину, которая затем выводится с мочой.
Печень делает из ненужных аминокислот нужные
Бывает, что в рационе человека не хватает каких-то аминокислот. Некоторые из них печень синтезирует, используя фрагменты других аминокислот. Однако некоторые аминокислоты печень делать не умеет, они называются незаменимыми и человек получает их только с пищей.
Печень превращает глюкозу в гликоген, а гликоген в глюкозу
В сыворотке крови должна быть постоянная концентрация глюкозы (иначе говоря – сахара). Она служит основным источником энергии для клеток головного мозга, мышечных клеток и эритроцитов. Самый надежный способ обеспечить постоянное снабжение клеток глюкозой – запасти ее после еды, а потом использовать по мере необходимости. Эта важнейшая задача возложена на печень. Глюкоза растворима в воде, и запасать ее неудобно. Поэтому печень вылавливает из крови избыток молекул глюкозы и превращает в нерастворимый полисахарид гликоген, который откладывается в виде гранул в клетках печени, а при необходимости снова превращается в глюкозу и поступает в кровь. Запаса гликогена в печени хватает на 12-18 часов.
Печень запасает витамины и микроэлементы
Печень запасает жирорастворимые витамины А, D, Е и К, а также водорастворимые витамины С, В12, никотиновую и фолиевую кислоты. А еще этот орган хранит минеральные вещества, нужные организму в очень малых количествах, такие как медь, цинк, кобальт и молибден.
Печень разрушает старые эритроциты
У человеческого плода эритроциты (красные кровяные тельца, которые переносят кислород), образуются в печени. Постепенно эту функцию берут на себя клетки костного мозга, а печень начинает играть прямо противоположную роль – не создает эритроциты, а разрушает их. Эритроциты живут около 120 дней, а затем стареют и подлежат удалению из организма. В печени есть специальные клетки, которые отлавливают и разрушают старые эритроциты. При этом освобождается гемоглобин, который вне эритроцитов организму не нужен. Гепатоциты разбирают гемоглобин на «запчасти»: аминокислоты, железо и зеленый пигмент. Железо печень хранит, пока оно не потребуется для образования новых эритроцитов в костном мозге, а зеленый пигмент превращает в желтый – билирубин. Билирубин поступает в кишечник вместе с желчью, которую окрашивает в желтый цвет. Если печень больна, билирубин накапливается в крови и окрашивает кожу – это желтуха.
Печень регулирует уровень некоторых гормонов и активных веществ
В этом органе переводится в неактивную форму или разрушается избыток гормонов. Их список довольно длинный, поэтому здесь мы упомянем только инсулин и глюкагон, которые участвуют в превращении глюкозы в гликоген, и половые гормоны тестостерон и эстрогены. При хронических болезнях печени метаболизм тестостерона и эстрогенов нарушен, и у пациента появляются сосудистые звездочки, выпадают волосы под мышками и на лобке, у мужчин атрофируются яички. Печень удаляет избыток таких активных веществ, как адреналин и брадикинин. Первый из них увеличивает частоту сердечных сокращений, уменьшает отток крови к внутренним органам, направляя ее к скелетным мышцам, стимулирует расщепление гликогена и повышение уровня глюкозы в крови, а второй регулирует водный и солевой баланс организма, сокращения гладкой мускулатуры и проницаемость капилляров, а также выполняет некоторые другие функции. Плохо бы нам пришлось при избытке брадикинина и адреналина.
Печень уничтожает микробов
В печени есть специальные клетки-макрофаги, которые располагаются вдоль кровеносных сосудов и вылавливают оттуда бактерии. Пойманные микроорганизмы эти клетки заглатывают и уничтожают.
Печень обезвреживает яды
Как мы уже поняли, печень – решительный противник всего лишнего в организме, и уж конечно она не потерпит в нем ядов и канцерогенных веществ. Обезвреживание ядов происходит в гепатоцитах. После сложных биохимических преобразований токсины превращаются в безвредные, растворимые в воде вещества, которые покидают наше тело с мочой или желчью. К сожалению, не все вещества удается обезвредить. Например, при распаде парацетамола образуется сильнодействующее вещество, которое может необратимо повредить печень. Если печень нездорова, или пациент принял слишком большую дозу парацетомола, последствия могут быть печальными, вплоть до гибели клеток печени.
По материалам zdorovie.info
Полезная информация, организации инвалидов, знакомства
источник
Сайт предоставляет справочную информацию исключительно для ознакомления. Диагностику и лечение заболеваний нужно проходить под наблюдением специалиста. У всех препаратов имеются противопоказания. Консультация специалиста обязательна!
Печень – это достаточно сложно устроенный орган. Морфологическая сложность строения тканей печени, разветвленная и замысловатая схема кровообращения и сеть желчных капилляров определяют многообразие функций данного органа. На самом деле печень выполняет ряд важнейших для нашего организма функций, каждая из которых жизненно необходима. Она является основным органом, осуществляющим обменные процессы организма, синтезирует ряд белков крови, осуществляет функцию обезвреживания токсичных веществ и их выведение, синтезирует желчь (тем самым активно участвуя в процессе кишечного пищеварения). На самом деле функций у печени намного больше, в данной статье мы коснемся лишь основных.
Как всем известно, печень является непарным органом, расположенном в правом подреберье. Обладая этими познаниями в анатомии, все, у кого закололо в правом боку, тот час диагностируют у себя заболевание печени. Это достаточно массивный орган, ее масса в среднем составляет 1,5 кг. Печень обладает отдельной сосудистой сетью, изолированной от общего кровотока. А причиной обособленной сосудистой сети является то обстоятельство, что в этот орган оттекает кровь от всего кишечного тракта. При этом печень является для крови, оттекающей от стенок кишечника естественным фильтром, исполняет функцию первичной сортировки, синтеза и распределения питательных веществ в организме. В кровеносную систему печени оттекает кровь практически от всех органов брюшной полости: кишечник (тонкий и толстый, желудок), селезенка, поджелудочная железа. Далее кровь, пройдя фильтрацию в тканях печени, вновь возвращается в большой круг кровообращения. Для того чтобы разобраться как функционирует печень, подробнее рассмотрим ее анатомическое и микроскопическое строение.
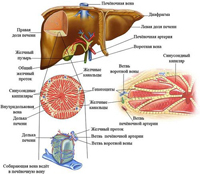 Морфологическое строение печеночной ткани достаточно сложное. Это высоко структурированная ткань с множеством особенностей. Но, как и все в живой природе в строении ткани печени главным является формула: «Функция определяет форму».
Морфологическое строение печеночной ткани достаточно сложное. Это высоко структурированная ткань с множеством особенностей. Но, как и все в живой природе в строении ткани печени главным является формула: «Функция определяет форму».
Итак, печень при ее рассмотрении под микроскопом имеет строение наподобие структуры пчелиных сот. Каждая печеночная долька имеет шестиугольную форму, в центре которой проходит центральная вена, а по периферии печеночная долька окутана сетью разнообразных сосудов: желчный проток, ветви воротной вены и печеночной артерии.
В просвете воротной вены кровь от органов брюшной полости движется по направлению к печеночным долькам.
По печеночной артерии происходит однонаправленный кровоток от сердца к тканям печени. Эта кровь обогащена питательными веществами и кислородом. Потому, основной функцией этой сети является обеспечение печеночной ткани энергетическими и строительными ресурсами.
По желчному протоку синтезированная гепатоцитами (клетки печени) желчь оттекает из печеночной дольки по направлению к желчному пузырю или просвету двенадцатиперстной кишки.
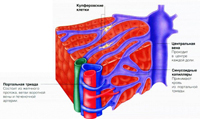 Напомним, что по воротной вене происходит приток крови к печени в основном от кишечника, со всеми растворенными в крови в результате пищеварения веществами. По печеночной артерии в печень поступает обогащенная кислородом и питательными веществами кровь от сердца. Внутри печеночной дольки сосуды, по которым поступает кровь в печеночную дольку, сливаются, образуя расширенную полость – синусоидные капилляры.
Напомним, что по воротной вене происходит приток крови к печени в основном от кишечника, со всеми растворенными в крови в результате пищеварения веществами. По печеночной артерии в печень поступает обогащенная кислородом и питательными веществами кровь от сердца. Внутри печеночной дольки сосуды, по которым поступает кровь в печеночную дольку, сливаются, образуя расширенную полость – синусоидные капилляры.
Проходя по синусоидным капиллярам, кровь значительно замедляет свою скорость. Это необходимо, для того чтобы гепатоциты успевали улавливать растворенные в крови вещества для их дальнейшей обработки. Питательные вещества подвергаются дальнейшей обработке и с током крови распространяются по сосудистой сети, или накапливаются в виде запасов в печени. Токсичные же вещества улавливаются гепатоцитами и обезвреживаются для последующего выведения из организма. Пойдя по синусоидным, капиллярам кровь попадает в центральную вену, расположенную в центре печеночной дольки. По печеночной вене кровь удаляется из печеночной дольки по направлению к сердцу.
Печеночные клетки выстроены в виде одноклеточных пластин расположенных перпендикулярно стенкам центральной вены. Внешне это напоминает развернутую на 360 градусов книгу, где торец – это центральная вена, а листы – трабекулы, между которыми переплетаются сосуды.
В отношении белков, жиров и углеводов важен тот факт, что эти вещества могут синтезироваться в печени. Причем углеводы могут синтезироваться из жиров или аминокислот. Жиры могут синтезироваться из продуктов расщепления углеводов и аминокислот. И только аминокислоты не могут быть синтезированы из углеводов или жиров. Витамины так же не синтезируются в нашем организме. Потому без постоянного поступления с пищей аминокислот и витаминов долго чувствовать себя здоровым невозможно.
Итак, в процессе пищеварения в крови, оттекающей от стенок кишечника множество расщепленных до уровня мельчайших жировых частиц (хиломикроны). В этой крови жиры, образуя эмульсию которая, по внешнему виду напоминает молоко. Углеводы, попадают в кровь в виде различных по строению молекул (фруктоза, мальтоза, галактоза и т.д.).
Аминокислоты – это структурные единицы белка, которые попадают в наш организм в виде отдельным молекул или в виде коротких цепочек скрепленных друг с другом частиц.
Аминокислоты – эти важные для нашего организма вещества с особой бережливостью используются клетками печени. Из них синтезируются ферменты, белки крови. Часть из синтезированных белковых молекул вновь возвращается в кровь для транспортировки к органам и тканям в виде аминокислот или белка плазмы крови – альбумина. Часть аминокислот расщепляется для построения других молекул аминокислот или иных органических веществ.
Витамины – эти вещества попадают в наш организм в процессе пищеварения, часть из них синтезируется микрофлорой кишечника. Однако все они поступают в организм, пройдя через печеночную ткань. Витамины являются незаменимыми веществами, поступающими в ткани печени с током крови. Витамины активно поглощаются клетками органа. Часть витаминов сразу встраивается в синтезируемые ферменты, часть запасается клетками печени, часть перенаправляется с током крови, оттекающим от данного органа к периферическим тканям. При прохождении печеночных синусов органические вещества и витамины улавливаются печеночными клетками и перемещаются внутрь гепатоцита. Далее, в зависимости от состояния организма происходят процессы преобразования и распределения.
Углеводы наиболее активно обрабатываются в печени. Многообразные формы углеводов преобразуются в единую – глюкозу. Далее глюкоза может высвобождаться в кровоток и по центральной вене устремляться в большой круг кровообращения, идти на энергетические нужды печени, либо расщепляться для производства необходимых организму веществ или накапливаться в виде гликогена.
Жиры – поступают в печень в виде эмульсии. При попадании в гепатоцит происходит их расщепление, жиры расщепляются на составные части глицерин и жирные кислоты. В дальнейшем из вновь синтезированных жиров формируются транспортные формы — липопротеиды из молекул холестерина, липидов и белка. Именно эти липопротеиды, поступая в кровоток, доставляют к периферическим тканям и органам жиры холестерин.
Сборка некоторых необходимых организму веществ осуществляется непосредственно в печени. И она обеспечивает не только трансформацию органических веществ и формирование их транспортных форм, но и синтезирует конечные формы белков, которые активно участвуют в обменных процессах, обеспечивают свертываемость крови, перенос некоторых гормонов и поддержание онкотического давления. Остановим свое внимание на некоторых из них:
Альбумин – это низкомолекулярный белок с молекулярной массой в 65000. Синтезируется сывороточный альбумин исключительно печенью. Количество альбумина содержащегося в литре сыворотки крови достигает 35 — 50 грамм. Альбумин осуществляет множество функций крови: является одной из транспортных форм белка в организме, осуществляет перенос на своей поверхности некоторых гормонов, органических веществ и медикаментозных препаратов, обеспечивает онкотическое давление крови (это давление препятствует выходу жидкой части крови за пределы сосудистого русла).
Фибрин – это низкомолекулярный белок крови, образующийся в печени благодаря ферментной обработке и обеспечивающий свертывание крои и образования тромба.
Гликоген – это молекулярное соединение, которое объединяет в виде цепочки молекулы углеводов. Гликоген исполняет функцию депо углеводов печени. В случае необходимости в энергетических ресурсах происходит расщепление гликогена и высвобождение глюкозы.
Печень – это орган, в котором имеется постоянная высокая концентрация основных структурных элементов: белки, жиры, углеводы. Для их транспортировки или хранения в тканях данного органа необходимо синтезирование более сложных молекул. Часть синтезированных молекул и микроскопических структур являются лишь транспортными формами белков (альбумин, аминокислоты, полипептиды), жиров (липопротеиды низкой плотности), углеводов (глюкоза).
Желчь – это биологическая жидкость коричневато-зеленого цвета, имеющая сложный состав. Вырабатывается она клетками печени (гепатоциты). Состав желчи сложен и представлен желчными кислотами, пигментными кислотами, холестерином и сложными жирами. Синтезируясь в печеночных дольках, желчь направляется из печени по желчевыводящим путям оп направлению к просвету кишечника. Она может либо направляться непосредственно в просвет двенадцатиперстной кишки или накапливаться в резервуаре – в желчном пузыре. Желчные кислоты, в просвете кишечника активно воздействуют на жиры, преобразуя последние в мелкодисперсную систему (измельчая большие капли жира до более мелких, вплоть до формирования жировой эмульсии). Именно благодаря желчи становится возможным расщепление и усваивание жиров.
Автор: Ткач И.С. врач, хирург офтальмолог
источник
ЖИРОВОЙ ОБМЕН — совокупность процессов превращений нейтральных жиров (триглицеридов) в организме животных и человека. К этим процессам относятся: 1) переваривание и всасывание жиров и продуктов их распада в жел.-киш. тракте; 2) промежуточный обмен жиров и жирных к-т в организме; 3) выделение жиров и продуктов их обмена из организма. Нарушения Ж. о. различной этиологии служат причиной развития многих патол. состояний.
В состав тканей животных и растений входят нейтральные жиры (см.) и жироподобные соединения (липоиды), объединенные общим названием липиды (см.), поэтому понятия «жировой обмен» и «липидный обмен» часто понимаются как синонимы.
Начало изучения Ж. о. относится к 1823 г., когда Шеврель (М. Chevreul) раскрыл хим. природу триглицеридов (см.), которые оказались сложными эфирами жирных кислот (см.) и глицерина (см.). Это открытие послужило стимулом к проведению исследований по изучению хим. состава различных жиров, в т. ч. жиров животного происхождения. В середине 19 в. было установлено наличие расщепляющего (липазного) действия желудочного и панкреатического соков на жиры. Позднее А. Н. Лебедев доказал возможность отложения в жировой ткани животных неизмененного чужеродного жира. В 1904 г. Ф. Кнооп создал оригинальную теорию окисления жирных кислот.
Значительные успехи в изучении обмена жиров и промежуточных продуктов их метаболизма, в первую очередь жирных к-т, были достигнуты во второй половине 20 в. в связи с широким использованием меченых атомов и общим прогрессом в развитии биохимии.
В организм взрослого человека с пищей ежесуточно поступает в среднем 70 г жиров животного и растительного происхождения. В полости рта жиры не подвергаются никаким изменениям, т. к. слюна не содержит расщепляющих жиры ферментов. Частичное расщепление жиров на глицерин (или моно- и диглицериды) и жирные к-ты начинается в желудке. Однако расщепление жиров в желудке протекает с небольшой скоростью, что объясняется следующими причинами. Во-первых, в желудочном соке взрослого человека и млекопитающих активность липазы (см.) крайне невысока. Во-вторых, величина pH желудочного сока далека от оптимальной для действия этого фермента (оптимальное значение pH для желудочной липазы находится в пределах 5,5—7,5). В-третьих, в желудке отсутствуют условия для эмульгирования жиров, а липаза может активно гидролизовать только жир, находящийся в форме жировой эмульсии. Поэтому у взрослых людей неэмульгированные жиры, составляющие основную массу пищевого жира, проходят через желудок без особых изменений.
Вместе с тем гидролитическое расщепление жиров в полости желудка играет важную роль в процессе пищеварения у детей, особенно у детей грудного возраста. Умеренная кислотность желудочного сока у детей грудного возраста (pH ок. 5,0) способствует перевариванию эмульгированного жира молока желудочной липазой. Кроме того, при длительном употреблении молока в качестве основного продукта питания возможно адаптивное усиление синтеза желудочной липазы у детей грудного возраста.
В 70-х гг. 20 в. Скау (R. Scow) было установлено, что слизистая оболочка верхней части пищевода грудного ребенка секретирует свою собственную липазу в ответ на сосательные движения ребенка при кормлении грудью. Возможно, что именно эта липаза и проявляет свое действие в желудке детей грудного возраста.
Несмотря на то, что расщепление жиров в желудке взрослого человека невелико, оно в какой-то степени облегчает последующее переваривание жиров в кишечнике. Во-первых, в желудке происходит частичное разрушение липопротеидных комплексов мембран клеток пищи, что делает жиры более доступными для последующего воздействия на них липазы панкреатического сока. Во-вторых, даже незначительное по объему расщепление жиров в желудке приводит к появлению свободных жирных к-т, которые, не подвергаясь всасыванию в желудке, поступают в кишечник и там способствуют эмульгированию жиров.
Основная часть пищевых жиров подвергается расщеплению в верхних отделах тонкого кишечника при действии липазы панкреатического сока. Панкреатическая липаза (КФ 3.1.1.3) проявляет оптимум действия при pH ок. 8,0 и расщепляет субстраты, находящиеся в эмульгированном состоянии (действие ее на растворенные субстраты значительно слабее). Фермент катализирует гидролиз эфирных связей в молекуле триглицерида, в альфа-положениях, в результате чего образуются две молекулы жирных к-т и бета-моноглицерид. В качестве промежуточных продуктов липолиза образуются альфа-, бета-диглицериды (схема 1):
В кишечном соке содержится липаза, катализирующая гидролитическое расщепление моноглицеридов и не действующая на ди- и триглицериды. Ее активность, однако, невысока, поэтому практически основными продуктами, образующимися в кишечнике при расщеплении пищевых жиров, являются жирные к-ты и бета-моноглицериды. Что касается глицерина, то при расщеплении триглицеридов в кишечнике он образуется в небольших количествах.
Эмульгирование жиров зависит от ряда факторов. Наиболее сильным эмульгирующим действием обладают желчные кислоты (см.), попадающие в двенадцатиперстную кишку с желчью (см.) в виде натриевых солей соответствующих конъюгатов, гл. обр. гликохолеата и таурохолеата. В двенадцатиперстную кишку вместе с пищевой массой заносится нек-рое количество желудочного сока, содержащего соляную к-ту, к-рая в двенадцатиперстной кишке нейтрализуется в основном бикарбонатами, содержащимися в панкреатическом и кишечном соках и желчи. Образующиеся при разложении бикарбонатов пузырьки углекислого газа разрыхляют пищевую кашицу и способствуют более полному перемешиванию ее с пищеварительными соками. Одновременно начинается эмульгирование жира. Соли желчных к-т адсорбируются в присутствии небольших количеств свободных жирных к-т и моноглицеридов на поверхности капелек жира в виде тончайшей пленки, препятствующей слиянию этих капелек. Кроме того, соли желчных к-т, уменьшая поверхностное натяжение на поверхности раздела двух фаз — воды и жира, способствуют расчленению больших капелек жира на меньшие. Создаются условия для образования тонкой и устойчивой жировой эмульсии с размером частиц 0,5 мкм и меньше. В результате эмульгирования резко увеличивается поверхность жиров, что облегчает взаимодействие их с липазой, т. е. ускоряет ферментативный гидролиз, а также всасывание (см.).
Всасывание жиров, как и других липидов (см.), происходит в проксимальной части тонкого кишечника. Тонко эмульгированные жиры могут частично всасываться через стенку кишечника без предварительного гидролиза. В этом случае фактором, лимитирующим этот процесс, по-видимому, является величина жировых капелек эмульсии, к-рая не должна превышать 0,5 мкм. Однако основная часть жира всасывается лишь после расщепления его панкреатической липазой на жирные к-ты и моноглицериды. Всасывание этих соединений происходит при участии желчи. В желчи содержатся соли желчных к-т, фосфатиды (см.) и холестерин примерно в соотношении 12,5 : 2,5 : 1,0. Жирные к-ты и моноглицериды образуют с этими компонентами желчи мицеллы, состоящие из солей желчных к-т, фосфатидов, холестерина, жирных к-т и моноглицеридов. Эти мицеллы могут связывать дополнительные количества неэтерифицированного холестерина, а также жирорастворимые витамины (A, D, Е и К). Структура мицелл такова, что их гидрофобное ядро, состоящее из липидов, оказывается окруженным снаружи гидрофильной оболочкой из желчных к-т и фосфатидов. Размеры мицелл примерно в 100 раз меньше самых мелких эмульгированных жировых капель. Благодаря таким мелким размерам и наличию наружной гидрофильной оболочки мицеллы в водной среде находятся в виде р-ра, называемого мицеллярным. Жировые мицеллы рассматриваются как своеобразный комплекс липидов и продуктов их распада в кишечнике с желчными к-тами и другими компонентами желчи, обеспечивающий растворимость липидов и продуктов их распада в водной среде (так наз. мицеллярная солюбилизация липидов) и их всасывание в кишечную стенку.
Относительно механизма всасывания жировых мицелл единой точки зрения нет. Признается возможность непосредственного проникновения мицелл в эпителиальные клетки ворсинок слизистой оболочки тонкого кишечника целой частицей путем так наз. мицеллярной диффузии без затраты энергии. В клетках ворсинок происходит распад мицеллярного комплекса, при этом желчные к-ты сразу же поступают в кровь и с током крови через систему воротной вены доставляются в печень, где снова поступают в желчь. С другой точки зрения, проникновение жировых мицелл внутрь эпителиальных клеток частично или полностью осуществляется путем пиноцитоза (см.). Допускается также возможность перехода только липидных ингредиентов (молекулярная диффузия липидов) из жировых мицелл внутрь эпителиальных клеток при соприкосновении мицелл с поверхностью слизистой оболочки ворсинок кишечника. При этом соли желчных к-т не проникают внутрь клеток, а остаются в просвете кишечника и подвергаются обратному всасыванию, гл. обр. в подвздошной кишке. Некоторые исследователи считают, что вместо мицеллярной диффузии может иметь место активная передача мицеллами липидов на акцепторы клеточных мембран ворсинок слизистой оболочки кишечника, а затем уже переход липидов внутрь клеток.
Каков бы ни был механизм всасывания липидов внутрь кишечной стенки, исключительно важное значение при этом имеет энтерогепатическая циркуляция желчных кислот, обеспечивающая всасывание больших количеств жирных к-т и моноглицеридов (50—70, а иногда и более 100 г) при относительно невысоком общем пуле желчных к-т (2,8—3,5 г).
Как уже упоминалось, при переваривании жиров образуются небольшие количества глицерина. Будучи хорошо растворимым в воде, глицерин легко всасывается в тонком кишечнике. Частично он подвергается превращению в альфа-глицерофосфат в эпителиальных клетках кишечника, частично поступает в кровяное русло. Жирные к-ты с короткой углеродной цепью (менее 10 С-атомов) также легко всасываются в кишечнике и поступают в кровь воротной вены, минуя какие-либо превращения в кишечной стенке.
Проникшие в цитоплазму эпителиальных клеток слизистой оболочки ворсинок тонкого кишечника свободные жирные к-ты и моноглицериды задерживаются в эндоплазматическом ретикулуме. Здесь из жирных к-т образуется их активная форма — ацил-КоА и происходит ацилирование моноглицеридов с образованием сначала диглицеридов, а затем триглицеридов. Т. о., продукты расщепления пищевых жиров, образовавшиеся в кишечнике и поступившие в его стенку, используются для ресинтеза жиров.
Биол, смысл этого процесса сводится к тому, что в стенке кишечника синтезируются жиры, специфичные для данного вида животного и отличающиеся по своей природе от пищевого жира. В известной степени это обеспечивается тем, что в синтезе триглицеридов и фосфатидов в кишечной стенке принимают участие наряду с экзогенными и эндогенные жирные к-ты, синтезированные заново или «модифицированные», напр, путем удлинения углеродной цепи, а также доставленные в клетки кишечной стенки из кровяного русла или синтезированные в печени и попавшие сначала в кишечник в составе фосфатидов желчи, а затем проникшие в эпителиальные клетки кишечника в составе липидных мицелл. Кроме того, в клетках кишечника происходит своеобразное перераспределение общего пула жирных к-т; напр., жирная к-та экзогенного триглицерид-ного происхождения может быть использована для этерификации холестерина как экзогенного, так и эндогенного происхождения; жирная к-та, ранее входившая в состав фосфатидов, может быть использована для ресинтеза триглицеридов и наоборот и т. д.
Наконец, в эпителиальных клетках тонкого кишечника содержатся ферменты — моноглицеридлипаза, расщепляющая моноглицерид на глицерин и жирную к-ту, и глицеролкиназа (КФ 2.7.1.30), катализирующая превращение глицерина, образовавшегося в ходе предыдущей реакции или всосавшегося из кишечника, в альфа-глицерофосфат. альфа-Глицерофосфат, взаимодействуя с жирнокислотным ацил-КоА, участвует в образовании фосфатидной к-ты, к-рая в стенке кишечника используется гл. обр. для ресинтеза фосфатидов, но частично может быть использована и для ресинтеза триглицеридов, в которых все три жирнокислотных остатка являются в таком случае заново синтезированными.
Однако способность организма к синтезу в клетках стенки кишечника жира, специфичного для данного вида животного, все же ограничена. А. Н. Лебедевым показано, напр., что при скармливании голодавшим собакам больших количеств чужеродного жира (напр., растительного масла) часть его обнаруживается в жировых тканях животного в неизмененном виде. Жировые депо, по-видимому, являются единственной тканью, где могут откладываться чужеродные жиры (этим пользуются в животноводстве при откорме на сало животных, напр, свиней; качество жира, образующегося при откорме, зависит от состава задаваемых кормов). Что касается жира, откладывающегося или вступающего в обмен в клетках других органов и тканей, то состав его отличается высокой специфичностью и мало зависит от пищевых жиров.
Механизм ресинтеза триглицеридов и фосфатидов в клетках стенки кишечника в общих чертах идентичен их биосинтезу в других тканях.
Ресинтезированные в клетках кишечной стенки триглицериды и фосфатиды, а также поступивший в эти клетки из полости кишечника холестерин, который частично там этерифицируется, соединяются внутри цистерн гладкого эндоплазматического ретикулума с небольшим количеством белка и образуют относительно стабильные комплексные частицы — хиломикроны. Хиломикроны содержат ок. 2% белка, 84—87% триглицеридов, 4—7% фосфатидов и от 2 до 5% холестерина. Размер частиц хиломикронов колеблется от 120 до 1100 нм. Благодаря большим размерам частиц хиломикроны не способны проникать из эндотелиальных клеток кишечника в кровеносные капилляры и диффундируют в лимф, систему кишечника, а из последней— в грудной лимф, проток. Из грудного лимф, протока хиломикроны поступают в кровь. Т. о., благодаря хиломикронам осуществляется транспорт экзогенных триглицеридов, холестерина и, частично, фосфатидов из кишечника через лимф, систему в кровь.
Через 2 часа после приема пищи, содержащей жиры, развивается так наз. алиментарная гиперлипемия (см. Липемия), характеризующаяся в первую очередь повышением концентрации триглицеридов в крови и появлением в ней хиломикронов. После приема слишком жирной пищи плазма крови принимает молочный цвет, что объясняется появлением в ней большого количества хиломикронов. Пик алиментарной гиперлипемии приходится на период между 4—6 час. после приема жирной пищи. Через 10— 12 час. содержание жира в сыворотке крови возвращается к норме. К этому же времени у здоровых людей из плазмы крови полностью исчезают хиломикроны. В связи с этим взятие крови для исследования содержания в ней липидов должно проводиться натощак спустя 14 час. после приема пищи.
Печень и жировая ткань играют наиболее важную роль в дальнейшей судьбе хиломикронов. Т. к. эндотелиальный слой синусоидов печени имеет большие отверстия, хиломикроны свободно диффундируют из плазмы крови в межклеточные пространства печени. По мнению одних исследователей, при контакте хиломикронов с оболочкой паренхиматозных клеток последние захватывают только триглицериды хиломикронов; по мнению других, в паренхиматозные клетки попадают продукты гидролиза триглицеридов хиломикронов — глицерин и жирные к-ты. Поэтому допускают, что гидролиз триглицеридов хиломикронов имеет место как внутри печеночных клеток, так и снаружи, на их поверхности.
В клетках печени имеются ферментные системы, катализирующие превращение глицерина в альфа-глицерофосфат, а неэтерифицированных жирных к-т (НЭЖК)— в соответствующие ацилы КоА. Последние либо окисляются в печени с выделением энергии, либо используются для синтеза триглицеридов и фосфатидов. Синтезированные триглицериды и частично фосфатиды используются для образования липопротеидов очень низкой плотности (пре-бета-липопротеидов), которые секретируются печенью и поступают в кровь. Липопротеиды очень низкой плотности являются главной транспортной формой эндогенных триглицеридов и переносят за сутки в организме человека от 25 до 50 г триглицеридов. Хиломикроны из-за больших размеров не способны проникать в клетки жировой ткани, поэтому триглицеридная часть хиломикронов подвергается гидролизу на поверхности эндотелия капилляров жировой ткани при действии фермента липопротеидлипазы. Локализация этого фермента тесно связана с поверхностью эндотелия капилляров. При действии липопротеидлипазы на триглицериды хиломикронов (а также на триглицериды пре-бета-лииопротеидов) происходит образование жирных к-т и глицерина. Часть жирных к-т проходит внутрь жировых клеток, а часть связывается с альбуминами сыворотки крови и уносится с ее током. С током крови покидают капиллярную сеть жировой ткани глицерин, а также частицы хиломикронов и пре-бета-липопротеидов, оставшиеся после расщепления их триглицеридного компонента и получившие название «ремнантов». Ремнанты задерживаются в печени и подвергаются распаду.
После проникновения в жировые клетки жирные к-ты превращаются в метаболически активные формы (ацил-КоА) и вступают в реакцию с альфа-глицерофосфатом, образующимся в жировой ткани из глюкозы. В результате этого взаимодействия ресинтезируются триглицериды, которые пополняют общий запас триглицеридов жировой ткани.
Расщепление триглицеридов хиломикронов в кровеносных капиллярах жировой ткани и печени приводит к фактическому исчезновению самих хиломикронов и сопровождается «просветлением» плазмы крови, т. е. она теряет молочный цвет. Просветление ускоряется гепарином. Предполагается, что гепарин активирует солюбилизацию липопротеидлипазы и ее действие. При некоторых заболеваниях отмечается пониженная активность липопротеидлипазы, в результате чего в крови постоянно определяются большие количества хиломикронов (хиломикронемия).
Промежуточный обмен жиров включает в себя следующие процессы: мобилизацию жирных к-т из жировых депо и их окисление (бета-окисление — см. Жирные кислоты), биосинтез жирных к-т и триглицеридов и превращение непредельных жирных к-т.
В жировой ткани человека содержится большое количество жира, преимущественно в виде триглицеридов, которые выполняют в обмене жиров такую же функцию, как гликоген печени в обмене углеводов. Эти соединения представляют собой запасные вещества, которые могут потребляться при голодании, физ. работе и других состояниях организма, требующих затраты энергии. Запасы этих веществ пополняются после потребления пищи, и они, т. о., являются своеобразным буфером в процессах накопления и использования энергии в организме.
Значение триглицеридов жировой ткани как энергетического резерва убедительно подтверждается следующими цифрами. Организм здорового человека содержит ок. 15 кг триглицеридов (140 000 ккал) и только 0,35 кг гликогена (1410 ккал). Гликоген, две трети запасов к-рого находится в мышцах, используется организмом при кратковременных и внезапно возникающих нагрузках. Что касается триглицеридов жировой ткани, то при средней энергетической потребности взрослого человека в 3500 ккал в сутки 15 кг жира теоретически достаточно, чтобы обеспечить энергией 40-дневную потребность организма.
Триглицериды жировой ткани подвергаются липолизу под действием липаз. В жировой ткани содержится несколько липаз, из которых наибольшее значение имеют так наз. гормоночувствительная липаза (триглицеридлипаза), диглицерид-липаза и моноглицеридлипаза. Гормоночувствительная липаза находится в жировой ткани в неактивной форме и активируется под влиянием циклического 3′,5′-АМФ (аденозинмонофосфата). Последний образуется из АТФ при участии аденилатциклазы. Воздействие гормона (или гормонов) направлено на первичный клеточный рецептор, который, связывая гормон, модифицирует свою структуру и в такой форме активирует пусковой фермент липолиза— аденилатциклазу. Образовавшийся в результате этих реакций циклич. 3′,5′-АМФ активирует фермент протеинкиназу (КФ 2.7.1.37), который в свою очередь активирует путем фосфорилирования липазу, превращая ее тем самым из неактивной формы в активную (схема 2).
При действии на триглицериды активированной таким сложным путем триглицеридлипазы, а затем при действии на промежуточные продукты липолиза ди- и моноглицерид-липаз образуются конечные продукты — глицерин и НЭЖК, которые из жировой ткани поступают в кровяное русло (глицерин — в свободном виде; НЭЖК — в виде комплекса с альбуминами плазмы крови). В действительности же судьба НЭЖК в самой жировой ткани во многом зависит от содержания в последней глюкозы или, более точно, от интенсивности протекающего в ней гликолиза (см.). Это объясняется тем, что образующийся в ходе гликолиза диоксиацетонфосфат частично восстанавливается при действии глицерофосфатдегидрогеназы в альфа-глицерофосфат, который в свою очередь вступает в реакцию с жирными к-тами, в результате чего снова образуются триглицериды.
Ресинтезированные триглицериды остаются в жировой ткани, способствуя, т. о., сохранению ее общих запасов. Такое состояние имеет место у сытых животных. При голодании, когда содержание глюкозы в жировой ткани понижено и образуются лишь незначительные количества альфа-глицерофосфата, освободившиеся в ходе липолиза НЭЖК не могут быть использованы жировой тканью для ресинтеза триглицеридов и поэтому они быстро покидают эту ткань. Т. о., активация гликолиза в жировой ткани является фактором, способствующим накоплению в ней триглицеридов, а угнетение гликолиза, наоборот, способствует их удалению.
Во всех случаях усиление липолиза в жировой ткани сопровождается нарастанием концентрации свободных жирных к-т в крови. Комплекс альбумин — свободные жирные к-ты содержит 99% белка и всего 1% жирных к-т. Несмотря на такое относительно низкое содержание жирных к-т в этом комплексе, их транспорт осуществляется в больших размерах: в организме человека за сутки переносится от 50 до 150 г жирных к-т. Это объясняется высокой скоростью обновления комплекса (период его полураспада ок. 5 мин.).
Связанные с альбуминами жирные к-ты попадают с током крови в органы и ткани, где подвергаются бета-окислению (см. Жирные кислоты), а затем окислению в цикле Трикарбоновых к-т (см. Трикарбоновых кислот цикл). Ок. 30% жирных к-т задерживается в печени уже при однократном прохождении через нее крови, где часть их используется для синтеза триглицеридов. Ресинтезированные триглицериды идут на образование пре-бета-липопротеидов. Нек-рое количество жирных к-т окисляется в печени до кетоновых тел (см.)— ацетоуксусной и бета-оксимасляной к-т. Кетоновые тела, не подвергаясь дальнейшим превращениям в печени, попадают с током крови в другие органы и ткани (мышцы, сердце и др.), где окисляются до CO2 и H2O. Небольшая часть мобилизованных жирных к-т используется в различных тканях для этерификации холестерина, синтеза фосфатидов, сфинголипидов и других соединений.
Триглицериды синтезируются во многих органах и тканях, но наиболее важную роль в этом отношении играют печень, стенка кишечника и жировая ткань. Путь биосинтеза триглицеридов протекает через образование альфа-глицерофосфата как промежуточного соединения.
В стенке кишечника и почках, где активность глицеролкиназы (КФ 2.7.1.30) высока, альфа-глицерофосфат образуется непосредственно путем фосфорилирования глицерина при участии АТФ. В жировой ткани и мышцах вследствие чрезвычайно низкой активности этого фермента путь образования альфа-глицерофосфата связан с гликолизом (или гликогенолизом). В печени имеют место оба пути образования альфа-глицерофосфата. альфа-Глицерофосфат реагирует с двумя молекулами активированных жирных к-т, в результате чего образуется фосфатидная к-та:
Фосфатидная к-та, одно время считавшаяся гипотетическим промежуточным соединением в биосинтезе триглицеридов и фосфатидов, обнаружена во многих тканях. В печени, напр., на ее долю приходится ок. 1% всех фосфатидов. Фосфатидная к-та при действии фосфатазы фосфатидной к-ты (фосфатидатфосфатазы; КФ 3.1.3.4.) теряет фосфорную к-ту и превращается в альфа,бета-диглицерид:
альфа,бета-Диглицерид является главным конечным субстратом в синтезе триглицеридов, а также фосфатидов в большинстве органов и тканей, за исключением кишечной стенки. Завершающая реакция протекает путем взаимодействия диглицерида с активированной жирной к-той (ацил-КоА):
В стенке кишечника, как отмечалось выше, для ресинтеза триглицеридов используются моноглицериды, в больших количествах поступающие из кишечника после расщепления пищевых жиров. При этом осуществляется следующая последовательность реакций: моноглицерид + жирнокислотный ацил-КоА -> диглицерид; диглицерид + жирнокислотный ацил-КоА —> триглицерид.
В норме количество нейтральных жиров и жирных к-т, выделяющихся из организма человека в неизмененном виде, не превышает 5% от количества жира, принятого с пищей. При этом основной путь выведения жира и жирных к-т осуществляется через кожу с секретами сальных и потовых желез. В секрете потовых желез содержатся гл. обр. водорастворимые жирные к-ты с короткой углеродной цепью; в секрете сальных желез преобладают нейтральные жиры, эфиры холестерина с высшими жирными к-тами и свободные высшие жирные к-ты, прогоркание которых обусловливает неприятный запах этих секретов. Небольшое количество жира выделяется в составе отторгающихся клеток эпидермиса.
При заболеваниях кожи, сопровождающихся усиленной секрецией сальных желез (себорея, псориаз, угри и др.) или усиленным ороговением и слущиванием эпителиальных клеток, выведение жира и жирных к-т через кожу значительно увеличивается.
В процессе переваривания жиров в жел.-киш. тракте всасыванию подвергается ок. 98% жирных к-т, входящих в состав пищевых жиров, и практически весь образовавшийся глицерин. Оставшаяся небольшая часть жирных к-т выделяется с фекалиями в неизмененном виде или жеподвергается воздействию микробной флоры кишечника. При этом происходит восстановление непредельных жирных к-т, образование оксикислот, к-т с разветвленной боковой цепью и т. п. В целом за сутки у человека с фекалиями выделяется ок. 5 г жирных к-т, причем не менее чем половина их имеет полностью микробное происхождение. С мочой выделяется небольшое количество короткоцепочечных жирных к-т (уксусная, масляная, валериановая), а также бета-оксимасляная и ацетоуксусная к-ты. Количество последних в суточной моче составляет от 3 до 15 мг. Появление высших жирных к-т в моче наблюдается при липоидном нефрозе, переломах трубчатых костей, при заболеваниях мочевых путей, сопровождающихся усиленным слущиванием эпителия, и при состояниях, связанных с появлением в моче альбумина (альбуминурия).
Ж. о. находится под регулирующим влиянием ц. н. с., к-рое осуществляется непосредственным направлением импульсов к жировым тканям или к железам внутренней секреции. Участие высших отделов ц. н. с. в регуляции Ж. о. наглядно иллюстрируется экспериментами, в которых повреждение гипоталамической области мозга вызывало у животных развитие ожирения.
Влияние ц. н. с., как было установлено еще исследованиями И. П. Павлова и его сотрудников, проявляется уже на этапе расщепления и всасывания жиров в жел.-киш. тракте путем воздействия на количество секретируемых пищеварительных соков и на содержание в них в зависимости от состава пищи различных ферментов, в т. ч. и липаз. Денервация изолированных оперативным путем участков жел.-киш. тракта, а также состояние наркоза приводят к замедлению расщепления и всасывания жиров.
Нейрогормональное влияние на Ж. о. в первую очередь связано с регулированием процесса мобилизации жирных к-т из жировых депо (табл.) Хорошо известно, что при эмоциональных стрессах в крови резко повышается содержание НЭЖК. Этот феномен объясняется резким увеличением выброса в кровь катехоламинов и активацией последними аденилатциклазы в жировой ткани. В конечном итоге происходит активация липолиза и освобождение НЭЖК. Длительно продолжающийся отрицательный эмоциональный стресс может вызвать таким путем заметное похудание. Жировая ткань, кроме того, иннервируется волокнами симпатической нервной системы, и возбуждение этих волокон сопровождается выделением норадреналина непосредственно в жировую ткань и активацией в ней липолиза. Вот почему денервация жировой ткани приводит к накоплению в ней жира. Через активацию или угнетение липолиза осуществляется действие на Ж. о. и многих других гормонов — глюкокортикоидов (см. Глюкокортикоидные гормоны), глюкагона (см.), АКТГ (см. Адренокортикотропный гормон), липомобилизующего фактора гипофиза (см. Липотропные факторы гипофиза), тироксина (см.), а также реализуется влияние различных физиол, состояний организма (голодание, охлаждение и др.).
Таблица. Влияние некоторых факторов на мобилизацию жирных кислот из жировой ткани
Предполагаемый механизм действия
Катехоламины, глюкагон, тироксин, глюкокортикоиды
Ускорение синтеза аденилатциклазы и гормоночувствительной липазы
Ослабление действия катехоламинов на аденилатциклазу; угнетение аденилатциклазы
Торможение освобождения жирных к-т в результате активации гликолиза в жировой ткани; угнетение аденилатциклазы
Стресс, физическая нагрузка, голодание, охлаждение
Стимуляция секреции катехоламинов и угнетение секреции инсулина
Как уже отмечалось, увеличение концентрации глюкозы в жировой ткани и повышение скорости гликолиза угнетают липолиз. К этому следует добавить, что повышение концентрации глюкозы в крови стимулирует секрецию инсулина, являющегося ингибитором аденилатциклазы, что также приводит к угнетению липолиза. Т. о., когда в организм поступает достаточное количество углеводов и скорость их расщепления высока, мобилизация НЭЖК и их окисление идут с пониженной скоростью. Как только запасы углеводов истощаются и снижается интенсивность гликолиза, происходит усиление липолиза, в результате чего ткани получают повышенные количества жирных к-т для окисления. Вместе с тем повышение содержания длинноцепочечных жирных к-т вызывает понижение интенсивности утилизации и окисления глюкозы, напр, в мышцах. Все это свидетельствует о том, что Ж. о. и углеводный обмен (см.)— главные энергообразующие процессы в животном организме — настолько тесно связаны друг с другом, что многие факторы, влияющие на один обмен, прямо или косвенно сказываются на течении другого.
Говоря о гормональном влиянии на Ж. о., следует отметить активирующее влияние гормона щитовидной железы на окисление жирных к-т, а также стимулирующее действие инсулина на синтез жиров из углеводов. Гиперфункция щитовидной железы сопровождается похуданием и уменьшением запасного жира, гипофункция — нередко ожирением. Удаление половых желез (кастрация) вызывает у животных и часто у человека избыточное отложение жира.
Немаловажное значение в регуляции Ж. о. имеет характер питания. Длительное избыточное потребление пищи, богатой жирами и углеводами, приводит к значительному отложению жира, и в этом случае никакие регуляторные механизмы не могут предотвратить этот процесс. При недостатке в пище липотропных веществ (см.), в частности фосфатидов или веществ, входящих в их состав (холина, инозита), а также метионина (см.) наблюдается избыточное отложение жира в печени («жировая печень»). Появление «жировой печени» объясняется скорее всего тем, что в отсутствие фосфатидов печень не может утилизировать триглицериды для образования липопротеидов. В поджелудочной железе обнаружено вещество липокаин (см.), введение к-рого предотвращает развитие «жировой печени».
Одной из причин пониженного всасывания жиров в тонком кишечнике может быть недостаточно полное их расщепление либо вследствие пониженной секреции панкреатического сока (недостаток панкреатической липазы), либо вследствие пониженного выделения желчи (недостаток желчных к-т для эмульгирования жира и образования жировых мицелл). Второй, наиболее частой причиной пониженного всасывания жира является нарушение функции кишечного эпителия, наблюдаемое при энтеритах, гиповитаминозах, недостаточности коры надпочечников и некоторых других патол, состояниях. В этом случае бета-моноглицериды и жирные к-ты, образовавшиеся в полости кишечника, не могут нормально всасываться из-за повреждения эпителиального покрова кишечника.
Нарушение всасывания жиров наблюдается при панкреатитах различной этиологии, закупорке желчевыводящих путей, массивной резекции тонкого кишечника, а также при осложнениях после хирургического вмешательства, при резекциях желудка и пищевода. В последнем случае причину нарушения всасывания жиров связывают с перерезкой волокон блуждающего нерва, приводящей к понижению тонуса желчного пузыря и замедленному поступлению желчи в кишечник. Пониженное всасывание жира отмечается при гипогаммаглобулинемиях, болезни спру (см.), болезни Уиппла (см. Интестинальная липодистрофия), при лучевой болезни, а иногда и при лучевой терапии.
Нарушение всасывания жира в тонком кишечнике приводит к появлению большого количества жира и жирных к-т в кале — стеаторее (см.). При длительном нарушении всасывания жира наблюдается пониженное поступление в организм жирорастворимых витаминов.
При недостаточной активности липопротеидлипазы наблюдается нарушение перехода жирных к-т из хиломикронов и липопротеидов низкой плотности (пре-бета-липопротеидов) плазмы крови в жировые депо. Наиболее резко это нарушение представлено при I типе гиперлипопротеинемии [по классификации Фредриксона (D. S. Fredrickson) с сотр.]. Этот тип гиперлипопротеинемии, называемый также «индуцированной жирами липемией» или гиперхиломикронемией, характеризуется тем, что вследствие полного отсутствия активности липопротеидлипазы наследственного характера триглицериды плазмы крови не могут подвергаться расщеплению и накапливаются в крови. Плазма крови в таких случаях имеет молочный цвет из-за чрезвычайно высокого содержания хиломикронов, а при ее стоянии всплывает сливкообразный слой хиломикронов. Наиболее эффективное лечение этого заболевания заключается в замене природных жиров синтетическими, содержащими короткоцепочечные жирные к-ты с 8—10 углеродными атомами, всасывающимися из кишечника непосредственно в кровь без предварительного образования хиломикронов. При V типе гиперлипопротеинемии, называемом также «смешанной гиперлипемией», вследствие пониженной активности липопротеидлипазы в крови накапливаются вместе с хиломикронами пре-бета-липопротеиды. Внутривенное введение гепарина, являющегося активатором липопротеидлипазы, больным с V типом гиперлипопротеинемии приводит к просветлению плазмы крови (при I типе введение гепарина не дает эффекта). При V типе гиперлипопротеинемии назначается диета с низким содержанием жиров и ограниченным содержанием углеводов.
При нарушении процесса превращения пре-бета-липопротеидов в бета-липопротеиды в крови накапливаются так наз. патологические «флотирующие» бета-липопротеиды (III тип гиперлипопротеинемии). Для больных с III типом гиперлипопротеинемии характерна пониженная толерантность к углеводам: нагрузка углеводами приводит к стойкому увеличению содержания триглицеридов и пре-бета-липопротеидов в крови. Характерно частое сочетание этого типа гиперлипопротеинемии с диабетом.
У практически здоровых людей, особенно среднего и пожилого возраста, нередко наблюдается избыточное отложение жира в подкожной жировой клетчатке и других жировых тканях. Причиной такого отложения жира является излишнее потребление пищи, общая калорийность к-рой превышает энергетические затраты организма. Избыточное отложение жира часто наблюдается при переходе от физически активной деятельности к малоподвижному образу жизни, когда сохраняется прежний уровень возбудимости пищевого центра и прежний аппетит, а энерготраты организма значительно снижаются. Патол. ожирение (см.) наблюдается в тех случаях, когда отдельно или в комплексе действуют следующие факторы: 1) пониженная активность жировой ткани в отношении мобилизации жира и повышенная активность ее в отношении отложения жира, 2) усиленный переход углеводов в жиры, 3) повышенная возбудимость пищевого центра, 4) пониженная по сравнению с нормой мышечная подвижность. Понижение активности жировой ткани в отношении мобилизации жира наблюдается, напр., при поражениях гипоталамических центров, когда ослабляются импульсы, идущие в жировую ткань по симпатическим путям и ускоряющие липолиз триглицеридов. Торможение мобилизации жира из его депо имеет также место при ослаблении функции щитовидной железы и гипофиза, гормоны которых (тироксин, тиреотропный и соматотропный гормоны, липомобилизующий фактор гипофиза) активируют липолиз. Пониженная функция половых желез приводит к избыточному отложению жира, особенно если она сопровождается нарушением деятельности гипофиза, гипоталамических центров и понижением мышечной активности (см. Адипозо-генитальная дистрофия; Иценко—Кушинга болезнь; Лоренса—Муна—Бидля синдром). Усиление перехода углеводов в жиры и отложение их в жировой ткани имеет место при повышенной секреции АКТГ, глюкокортикоидов и инсулина. Гиперсекреция инсулина является главным фактором в развитии так наз. наследственно-конституционного ожирения.
Необычное отложение жира может наблюдаться в отдельных участках жировой ткани, в т. ч. располагающихся по зонам распространения нервных волокон определенных сегментов ц. н. с., что указывает на то, что подобное отложение жира или, наоборот, атрофия подкожной жировой ткани связаны с изменением трофической функции нервной системы как в центральных его отделах, так и периферических (см. Деркума болезнь, Липоматоз, Липодистрофия, Маделунга синдром).
Недостаточное отложение жира в жировой ткани (исхудание) развивается вследствие угнетения возбудимости пищевого центра (см. Анорексия), понижения всасывания жиров и углеводов (напр., при энтеритах), преобладания процессов мобилизации жира из жировой ткани над его отложением и при длительном голодании.
Нарушение образования жира из углеводов может наблюдаться при поражениях вегетативных (трофических) центров гипофизарно-диэнцефалической системы, а также коры надпочечников. Такие нарушения лежат в основе прогрессирующего истощения при болезни Симмондса (см. Гипофизарная кахексия) и аддисоновой болезни (см.).
Накопление жира в печеночных паренхиматозных клетках часто является реакцией печени на различные заболевания, токсические воздействия и повреждения. С биохим, точки зрения накопление жира в печени происходит тогда, когда скорость образования в ней триглицеридов превышает скорость их утилизации (липолиз триглицеридов и последующее окисление жирных к-т, включение триглицеридов в пре-бета-липопротеиды и их секреция в кровяное русло).
Жировая инфильтрация печени наблюдается при диабете, ожирении, белковой недостаточности, при отравлении четыреххлористым углеродом, фосфором, алкоголем и при недостаточности липотропных веществ.
Одним из наиболее распространенных нарушений Ж. о. у человека является кетоз — повышенное образование в организме, накопление в тканях и крови и выделение с мочой кетоновых тел.
В результате радиоактивного облучения в различных тканях организма Ж. о. претерпевает выраженные изменения, которые имеют и качественные и временные особенности, зависящие от вида ткани. Поражение при облучении тонкого кишечника приводит к уменьшению содержания липопротеидов, фосфолипидов, жирных к-т и холестерина в слизистой оболочке, что обусловлено наряду с деструкцией клеток подавлением синтеза липидов. Активность гидролитических ферментов в слизистой оболочке тонкой кишки понижается, в результате чего снижается интенсивность расщепления жиров и всасывания триглицеридов; всасывание жирных к-т угнетается в меньшей степени. В зависимости от условий питания содержание липидов в печени увеличивается или остается нормальным, но в любом случае усиливается синтез общих липидов и их отдельных фракций — триглицеридов, фосфолипидов, жирных к-т и холестерина. При этом происходит угнетение синтеза жиров, сопряженного с гликолизом (или гликогенолизом) и активация синтеза жиров из уксусной к-ты.
В результате изменений Ж. о. после облучения в печени, как правило, развивается гиперлипемия (после неспецифической реакции). В ранние сроки после облучения гиперлипемия обусловлена увеличением содержания триглицеридов, фосфолипидов, холестерина и его эфиров, а в более поздние — и жирных к-т. В основе механизма пострадиационной гиперлипемии лежит не столько мобилизация липидов из периферических депо, сколько усиление их биосинтеза в печени. Механизм переноса липидов из крови в ткани после облучения не изменяется. Стимуляция биосинтеза жиров в печени из уксусной к-ты не является результатом прямого воздействия облучения на печень, а первично обусловлена пострадиационной деструкцией радиочувствительных тканей, катаболиты к-рой, гл. обр. лактат и аминокислоты, поступая в избытке в печень, индуцируют глюконеогенез, что сопряжено с угнетением гликолиза и соответственно синтеза липидов из глюкозы и усилением синтеза липидов из уксусной к-ты в результате активизации ферментной системы, катализирующей биосинтез жирных к-т.
В первые дни после облучения в жировой ткани усиливается синтез липидов из глюкозы и ее метаболитов, что также связано с активизацией глюконеогенеза и последующей гипергликемией и увеличением содержания инсулина в крови. В более поздние сроки после облучения синтез жиров в жировой ткани понижается и сменяется мобилизацией жиров. Указанные изменения Ж. о. начинают развиваться через несколько часов после облучения, вскоре достигают максимума и остаются на этом уровне в течение нескольких дней. У большинства экспериментальных животных эти нарушения Ж. о., вызванные облучением, нивелируются в течение недели, за исключением мобилизации липидов из депо, к-рая продолжается еще нек-рое время, и имеют количественные и качественные видовые особенности. В лимфоидной ткани (тимус и селезенка) через несколько часов после облучения увеличивается концентрация общих липидов и жирных к-т, а также стимулируется синтез холестерина и общих липидов. В эти же сроки в костном мозге наблюдается увеличение содержания жирных к-т, содержание же липидов возрастает позднее; на протяжении недели после облучения в костном мозге происходит синусоидальное увеличение и уменьшение концентрации насыщенных и ненасыщенных жирных к-т при нормальном общем содержании жирных к-т в сыворотке крови. Увеличение содержания липидов в костном мозге обусловлено усилением их биосинтеза преимущественно в кроветворных, а не в жировых клетках. Наряду с повышением интенсивности синтеза липидов в костном мозге после первоначального усиления отмечается резкое и длительное торможение окисления жирных к-т до CO2 и H2O, к-рое продолжается и после восстановления нормального клеточного состава костного мозга. В радиорезистентных тканях (головной мозг, почки, легкие, скелетные мышцы и мышцы сердца) существенных изменений в Ж. о. не происходит. Активизация синтеза холестерина в надпочечниках связана с развивающейся после облучения стрессовой реакцией и усилением биосинтеза кортикостероидов из холестерина.
Состав триглицеридов в жировой ткани новорожденного отличен от материнского, т. к. человеческий плод синтезирует индивидуальноспецифические жиры из НЭЖК и глюкозы, поступающих трансплацентарно или с током пуповинной крови.
Доля жира в теле новорожденного зависит от его веса при рождении: при весе 1500 г она составляет примерно 3%, 2500 г — 8%, а 3500 г — 16% (норма). Жировая ткань ребенка содержит большее по сравнению с жировой тканью взрослого человека количество ди-глицеридов, воды, гликогена, ДНК, фосфолипидов и меньшее — нейтральных жиров, ненасыщенных жирных к-т с длинной углеродной цепью, холестерина; характерно также снижение величины коэффициента холестерин/фосфолипиды. С возрастом эти особенности постепенно нивелируются.
Гистол, и анатомической особенностью жировой ткани новорожденных является наличие скоплений бурого жира (до 8% от веса тела) вокруг органов, между лопатками, позади грудины, вокруг шеи, вдоль позвоночника. Этот жир играет роль своеобразного терморегулятора в силу высокой теплопродукции, обеспечиваемой большим содержанием в митохондриях цитохромов. Гипоксемические и железодефицитные состояния у детей (пневмонии, анемии, интоксикации) сопровождаются истощением бурого жира, резким снижением теплопродукции, что может способствовать легкому возникновению переохлаждения новорожденного.
У ребенка всех возрастов выше, чем у взрослых, способность синтезировать и накапливать жир, интенсивнее окисление жирных к-т, активнее процесс включения 14C-глюкозы в жирные к-ты и триглицериды клеток жировой ткани, более высока концентрация КоА. По мере роста и развития ребенка эти различия стираются.
Липолитическая активность сыворотки крови у детей стабильна, мало отличается от активности липаз в сыворотке крови взрослых, но степень торможения липолиза глюкозой с возрастом увеличивается, а активация адреналином уменьшается.
В первые часы жизни ребенка наблюдаются значительные изменения величины соотношения НЭЖК и глицерина сыворотки крови; у грудного ребенка она ниже, чем у взрослых. При недоношенности и гипотрофии величина этого соотношения резко уменьшается; в крови снижается и содержание высших ненасыщенных жирных к-т относительно общего количества ненасыщенных жирных к-т, что объясняется замедленностью обменных процессов.
В крови новорожденных сравнительно низка концентрация липидов и липопротеидов. Так, среднее содержание общего холестерина составляет ок. 70 мг%, а триглицеридов ок. 40 мг%. При низком содержании всех классов липопротеидов характерно сравнительно высокое относительное содержание альфа-липопротеидов. Концентрация НЭЖК в крови новорожденных может быть ниже или выше по сравнению с концентрацией НЭЖК в крови матери,— это зависит от веса новорожденного, его доношенности или недоношенности, состояния питания (нормо-, гипо- или паратрофия); она подвержена более значительным, чем у взрослых, колебаниям под влиянием количества и качества пищи (углеводистая и жирная пища повышает концентрацию НЭЖК, белковая — снижает) и различных заболеваний. Физ. и психические нагрузки, голодание, гормоны коры надпочечников, гипофиза, щитовидной железы способствуют более легкому повышению концентрации НЭЖК, а введение глюкозы, инсулина и белка — такому же снижению концентрации НЭЖК в крови у детей (по сравнению со взрослыми).
У новорожденных детей отмечается преобладание НЭЖК над этерифицированными жирными к-тами (ЭЖК), у грудных детей и детей более старшего возраста это соотношение обратное, как и у взрослых; остальные показатели липидного обмена также приближаются к аналогичным показателям у взрослых.
У новорожденных детей и детей раннего возраста расщепление и всасывание жиров имеет ряд особенностей. Они обусловлены функциональной незрелостью печеночных клеток и проявляются относительным недостатком желчных к-т, лимитированным вследствие этого эмульгированием жиров в кишечнике и нарушением их переноса через эпителий слизистой оболочки. Поэтому у детей чаще, чем у взрослых, встречаются диареи, связанные с расстройством переваривания и всасывания липидов.
Концентрация хиломикронов нарастает пропорционально степени зрелости ребенка: у недоношенных их число в 1 мм2 счетной камеры составляет 1400, у доношенных— 7800. Это тоже является причиной более легкого развития нарушений липидного обмена у недоношенных детей.
Лабильность функционального состояния систем регуляции Ж. о., свойственная ребенку, ведет (особенно у недоношенных детей и у детей с гипотрофиями или паратрофиями) к более стойким и выраженным, чем у взрослых, изменениям состава липидов крови (дислипидемии) при воспалительных и аллергических заболеваниях кожи, инфекциях, интоксикациях, расстройствах пищеварения, повышенной чувствительности к аллергенам, при нарушениях роста и развития. У детей чаще наблюдается гиперлипемия, обусловленная адаптационным усилением мобилизации депонированных жиров, к-рое у недоношенных детей и детей с гипотрофиями вызывает такое быстрое и глубокое истощение энергоресурсов, что это может угрожать жизни больного. Эти особенности должны учитываться при назначении таким детям комплексной терапии (искусственное насыщение питательными веществами, смесями, близкими по составу к грудному молоку, глюкозой и физиологически активными препаратами: АТФ, витамином С и витаминами группы В, ферментами, ускоряющими переваривание и всасывание жиров и углеводов, анаболическими гормонами).
Не все нарушения липидного состава крови у детей носят вторичный характер. С целью ранней диагностики возможных первичных наследственных дислипидемий у ребенка всякая гиперлипемия и другие выявленные изменения Ж. о. требуют тщательного анализа этиол, факторов, учета семейного анамнеза, точной идентификации характера изменений, сопоставления их с клиникой и другими обменными и гормональными показателями. См. также Гаргоилизм, Гиперхолестеринемия, Гоше болезнь, Липемия, Липидозы, Ниманна-Пика болезнь, Сфинголипидозы, Хенда-Шюллера-Крисчена болезнь.
Нарушения Ж. о. при старении проявляются прежде всего в повышении общего содержания жира в организме. В старости снижается абсолютная скорость накопления жира, но относительный его прирост (в процентах к сухому весу) выше, чем в молодом возрасте. Общее содержание жира увеличивается и в тканях (в жировых депо, печени, скелетной мускулатуре, селезенке и др.), и в крови (у мужчин 60— 74 лет до 472,5 ± 27,6 мг%, у женщин этого же возраста до 517,9 ± 26,6 мг%). В старости, по данным большинства исследователей, увеличивается содержание общего холестерина в крови (см. Гиперхолестеринемия), максимальная его концентрация в крови (260 мг%) наблюдается у мужчин в 40—50 лет, а у женщин приблизительно на 10 лет позже. Повышение концентрации общего холестерина происходит как за счет его свободной, так и за счет эфирносвязанной фракций, содержание его увеличивается в стенках аорты, в межреберных и межпозвоночных хрящах, в головном мозге, печени, почках, селезенке, в сердечной и скелетных мышцах. В коже концентрация холестерина снижается, а в легких и надпочечниках остается на прежнем уровне. Не установлена прямая зависимость между концентрацией холестерина в крови и содержанием его в тканях. С возрастом холестерин накапливается и в субклеточных структурах (ядрах, митохондриях, микросомах), что может приводить к снижению проницаемости клеточных мембран и тем самым — к нарушению процессов обмена веществ.
С возрастом изменяется также и качественный состав фосфолипидов: в мозге повышается концентрация дифосфоинозитида, кефалина и особенно сфингомиелина. Количество фосфолипидов в сыворотке крови пожилых и старых людей достигает у мужчин 216,4 ± 3,9 мг% , у женщин 236,7 ± 7,9 мг%. Уменьшение величины отношения фосфолипиды/холестерин свидетельствует о снижении с возрастом надежности систем, регулирующих постоянную концентрацию холестерина в крови. В процессе старения в крови значительно увеличивается содержание триглицеридов: у старых мужчин оно возрастает до 113,9 ± 5,8 мг%, у женщин до 104,1 ± 6,0 мг%. При нагрузках жиром концентрация триглицеридов в крови старых людей повышается больше, чем у молодых, и медленнее возвращается к исходным величинам. Изучение скорости липогенеза по включению ацетата-1-14C свидетельствует о снижении при старении скорости синтеза триглицеридов как de novo, так и из преформированных жирных к-т. Повышение концентрации триглицеридов в крови в старости может быть обусловлено уменьшением скорости их распада, а также ухудшением транспорта триглицеридов из крови в ткани. При старении в крови повышается содержание НЭЖК, что, возможно, связано со снижением способности стареющего организма к их утилизации, в частности к их окислению в тканях. В старости увеличивается содержание в крови ненасыщенных жирных к-т, а также жирных к-т с длинной углеродной цепью. Это ведет к снижению проницаемости клеточных мембран и способствует образованию свободных радикалов, оказывающих повреждающее действие на генетический аппарат клетки и тем самым ускоряющих темп старения. С возрастом в крови людей повышается содержание а- и, главное, пре-бета-липопротеидов. Неравномерное изменение количества общего холестерина и холестерина, связанного с p-липопротеидами, свидетельствует о его перераспределении в течение жизни человека между липопротеидными фракциями. С возрастом увеличивается содержание атерогенных липопротеидов, растет число лиц, у которых обнаруживается гиперлипопротеинемия различной степени. Накопление холестерина в p-липопротеидах снижает стабильность липопротеидных комплексов и способствует отложению его в стенках артерий. С возрастом снижается и липолитическая активность в стенке аорты, в селезенке, легких, клапанах сердца. Изменения в Ж. о. при старении являются результатом сдвигов в различных звеньях его нейрогуморальной регуляции, ослабления влияния высших вегетативных центров на эндокринные железы, снижения активности последних, повышения чувствительности тканей к действию гормонов. Нарушение Ж. о. в старости создает метаболические предпосылки для развития ряда заболеваний — атеросклероза (см.), гипертонической болезни (см.), коронарной недостаточности, диабета и др.
Библиография: Алимова Е. К., Аствацатурьян А. Т. и Жаров Л. В. Липиды и жирные кислоты в норме и при ряде патологических состояний, М., 1975, библиогр.; Верещагин А. Г. Биохимия триглицеридов, М., 1972, библиогр.; Габуния Р. И. Метод радиометрии всего тела в клинической диагностике, М., 1975, библиогр.; Геронтология и гериатрия, 1971, Ежегодник, под ред. Д. Ф. Чеботарева, с. 82, Киев, 1971; Климов А. Н. и Никульчева Н.Г.Типы гиперлипопротеинемий, их связь с атеросклерозом и лечение, Кардиология, т. 12, № 6, с. 133, 1972, библиогр.; К о-р о в и н а Н. А., Барышков Ю. А. и Вельтищев Ю.Е. О физиологии и патологии липидного обмена у детей, Педиатрия, № 2, с. 74, 1967, библиогр.; Лейтес G. М. Физиология и патология жировой ткани, М., 1954; Липиды — структура, биосинтез, превращения и функции, под ред. G. Е. Северина, М., 1977; Мороз Б.Б. и К ендыш И. Н. Радиобиологический эффект и эндокринные факторы, М., 1975, библиогр.; Ныос-холм Э. и Старт К. Регуляция метаболизма, пер. с англ., с. 346, М., 1977; Рачев Л., Тодоров Й. и Статева Ст. Обмен веществ в детском возрасте, пер. с болг., София, 1967, библиогр.; Экспериментальный атеросклероз и возраст, под ред. H. Н. Горева, М., 1972, библиогр.; Adipose tissue, ed. by B. Jeanrenaud a. D. Hepp, Stuttgart a. o., 1970; Biochemical problems of lipids, ed. by A. C. Frazer, Amsterdam a. o., 1963; Blood lipids and lipoproteins, quantitation, composition and metabolism, ed. by G. J. Nelson, N. Y. a. o., 1972; Current trends in the biochemistry of lipids, International symposium on lipids, ed. by J. Ganguly a. R. M. S. Smellie, L.—N. Y., 1972; Intestinal absorption in man, ed. by I. Me Coli a. G. E. G. Sladen, L., 1975; Jackson R. L., Morrisett J. D. a. G o t t o A. M. Lipoprotein structure and metabolism, Physiol. Rev., v. 56, p. 259, 1976, bibliogr.; KingK. C, a. o. Sources of fatty acid in the newborn, Pediatrics, v. 47, Suppl. 2, p. 192, 1971; Lipid metabolism, ed. bv S. J. Wakil, N. Y.—L., 1970; Masoro E. J. Physiological chemistry of lipids in mammals, Philadelphia a. o., 1968; Scow R. O., В 1 a n c h e t t e — M a c k i e E. J. a. Smith L. C. Role of capillary endothelium in the clearance of chylomicrons, Circulat. Res., v. 39, p. 149, 1976; S e a r-c у R. L. Lipopathies, Springfield, 1971.
A. H. Климов; Л. H. Богацкая (геронт.), А. В. Картелишев (пед.), И. Н. Кендыш (рад. био.).
источник